








Services on Demand
Article
 Article in pdf format
Article in pdf format Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
 Cited by SciELO
Cited by SciELO
 Similars in SciELO
Similars in SciELO
Bookmark
Revista Científica Agrociencias Amazonía
Print version ISSN 2307-9606
Rev.Cient.Agro.Amaz. vol.1 no.2 Beni Nov. 2013
ARTICULO
Condiciones Opticas y Comunidades IctÃcolas en Aguas Blancas y Negras, RÃos Tahuamanu, Manuripi y Madre de Dios, AmazonÃa Boliviana
Yunoki T.1, Torres L.V.1 y Yagami T.2
1 Centro de Investigación de Recursos Acuáticos (CIRA), Facultad de Ciencias Pecuarias, Universidad Autónoma del Beni 'José Ballivián', Trinidad, Beni, Bolivia. Correo de contacto: takayukiyunoki@yahoo.com
2 Ex-Japanese Overseas Cooperation of Volunteer, Japan International Cooperation Agency, Civil Engineering and Eco-technology Consultants Co., LTD. Ecological Analysis Section, ZIP: 170-0013, 2-23-2 Higashiikebukuro, Toshimaku, Tokyo, Japan.
RESUMEN
Se describen las estructuras de comunidades ictÃcolas y la adaptabilidad a la condición óptica, mediante el modelo PiscÃvoro de Transparencia Morfológica (PTM), desde la perspectiva de la teorÃa de metacomunidad en los rÃos y lagunas de aguas blancas y negras en la AmazonÃa boliviana. Los tipos de agua cambian, indicando un posible patrón geológico o vegetal de origen entre los valles de aguas blancas y negras, asà mismo un patrón geomorfológico entre los rÃos y lagunas en los valles de aguas blancas. Las comunidades ictÃcolas cambian espacialmente junto con los tipos de aguas. El modelo PTM presenta mayor ajuste para especies restringidas y menor ajuste para especies con amplia distribución, indicando un balance de tácticas entre la adaptación a la condición óptica y la dispersión. Los taxones superiores con mayor diversidad poseen especies con mayor rango de distribución y menor control filogenético, las cuales son especies dominantes a escala regional. Estas especies al disminuir la adaptación a la condición óptica e incrementar la dispersión, han adquirido probablemente una ventaja comparativa en la cadena trófica. La aplicabilidad del modelo PTM desde la perspectiva de la teorÃa de meta comunidad, sugiere que los efectos de predadores sobre poblaciones de presas no solo son mortalidades directas causadas por éstos, sino son también consecuencia del comportamiento contra - predador, asà como de historias vitales por las cuales las comunidades ictÃcolas de diferentes tipos de aguas pueden coexistir y coevolucionar.
Key words: PTM model, metacommunity, adaptation
ABSTRACT
The fish communities structures and the optic condition adaptation are described, through the piscivory-transparency-morphometry (PTM) model from the perspective of metacommunity theory in the rivers and lagoons of the white and black water in Bolivia Amazon. The water types changed, indicating a possible geologic or vegetation origin pattern between white and black water valleys, also a geomorphologic pattern between the rivers and lagoons of white water valleys. Fish communities changed spatially according to the water types. The PTM model shows a better fit for species with small distribution and lesser fit for species with large distribution, indicating a balance between adaptation to the optic condition and dispersion. The higher taxas with higher diversity have the species of larger distribution and lesser phylogenetic control which are the dominant species in regional scale. These species having a largedistribution,probablyhavegainedadvantages in the trophic network. The applicability of the PTM model from the perspective of the metacommunity theory, suggests that the effects of predators on preys are not only the direct mortalities caused by them, but also the consequences of contra predator behaviors and the life history so that fish communities of different water types can coexist and coevolute.
Palabras claves: modelo PTM, metacomunidad, adaptación Yunoki T., Torres L.V.y Yagami T.
INTRODUCCIÃN
Las tierras bajas de la Amazonia están circunscritas por dos formaciones geológicas: Andes y Precámbrica. Sus valles están clasificados en tres categorÃas: valles de aguas blancas, claras y negras, reflejando la geologÃa y la vegetación de sus orÃgenes. Las caracterÃsticas geomorfológicas forman los hábitats como; varzea (zonas de inundación de valles de aguas blancas), tierra firme (terrazas), e igapo (zonas de inundación de valles de aguas negras). Las caracterÃsticas topográficas y el balance hÃdrico en varzea determinan la influencia de diferentes tipos de aguas lénticas y lóticas y las correspondientes comunidades de organismos acuáticos. Las lagunas de varzea donde las aguas blancas ricas en nutrientes decantan sus sedimentos son consideradas como más productivas (Fittkau et al., 1975).
En las lagunas de aguas blancas y negras del rÃo Orinoco venezolano, las estructuras de comunidades ictÃcolas están organizadas por los ictiófagos, bajo la influencia de transparencia controlada por profundidad y área (modelo PTM). Las lagunas claras están dominadas por los peces orientados por su visión; los Characiformes y los Cichlidae, mientras las lagunas turbias por los peces adaptados a la baja visibilidad; los Gymnotiformes y los Siluriformes (RodrÃguez y Lewis, 1997). Según estos autores, el modelo PTM es probablemente aplicable a otras zonas de inundación tropical. Las aguas blancas son más turbias, ricas en nutrientes y menos ácidas que aguas claras y negras.
Los estudios ecológicos han destacado los efectos quÃmicos de aguas sobre la productividad para interpretar el notorio cambio faunÃstico entre estos tipos de aguas, sin embargo el modelo PTM podrÃa ofrecer una descripción más precisa. Asà tambien, pretenden probar el efecto de factores bióticos, abióticos y espaciales sobre las estructuras de comunidades ictÃcolas. Por ejemplo, la depredación por peces es un factor determinante en el sistema fluvial. En los pequeños rÃos y lagunas, la depredación puede excluir otras especies. En una región geográfica, los factores abióticos frecuentemente determinan la importancia relativa de depredación, influyendo indirectamente en la composición de presas. La escala espacial de estudios influye la importancia percibida del factor biótico respecto al abiótico (Jackson etal., 2001). La teorÃa de metacomunidad, pretende ampliar la interacción biótica de la escala local en la comunidad homogénea y aislada, hacia la escala regional entre las comunidades discretas. La metacomunidad se define como un conjunto de comunidades interconectadas por la dispersión de múltiples especies que potencialmente interac-túan. Por el contrario, en la comunidad local, las especies interactúan entre ellas afectando sus tasas demográficas. Esto supone que existe por lo menos dos comunidades discretas y que, a escala regional, la dispersión entre comunidades locales ocurre a diferentes tasas (Leibold et al., 2004). La teorÃa de metacomunidad dirige el balance entre la dispersión, la heterogeneidad ambiental y la interacción biótica, para estructurar comunidades a lo largo de escalas espaciales, y está actualmente en el estado de infancia, siendo más heurÃstica que predictiva (Falke & Fausch, 2010).
En las tierras bajas de la Amazonia, la correspondencia entre tipos de aguas y comunidades ictÃcolas discretas fue denominadafrecuentemente como un evento biogeográfico y no fueron consideradas con mayor profundidad (Saint-Paul et al., 2000). El presente estudio describe las estructuras de comunidades ictÃcolas, y prueba el modelo PTM entre las comunidades locales discretas desde el punto de vista de la teorÃa de metacomunidad, analizando el balance de tácticas entre la adaptabilidad a la condición óptica y la dispersión en los rÃos y lagunas de aguas blancas y negras de los rÃos Tahuamanu, Manuripi y Madre de Dios en la AmazonÃa boliviana.
MATERIALES Y MÃTODOS
Ãrea de estudio. La zona estudiada está ubicada geográficamente entre el paralelo 11°00' y 13°00' de latitud Sur y 67°30' y 69°30' de longitud Occidental, correspondiendo al área Norte de la cuenca Amazonica boliviana. Abarca dos valles de aguas barrosas (Tahuamanu y Madre de Dios) y un valle de aguas oscuras (Manuripi), delimitados por terrazas (IGM, Mapa geográfico, 1:250 000, SC 19-14, SC 19-15, y SC 19-13, Servicio Nacional de HidrografÃa Naval, 1998) (Figura 1).
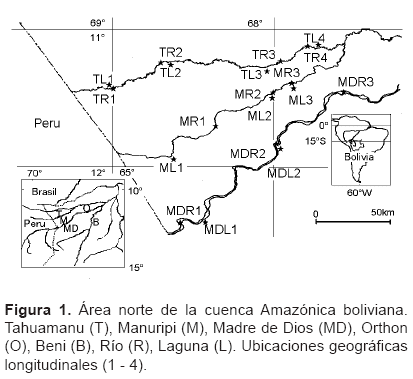
El área de estudio pertenece a un mismo régimen pluviométrico. La estación seca es en invierno, con un mÃnimo de mayo a julio (Roche et al., 1992). Durante la estación seca, la mayor parte de las lagunas de los rÃos Tahuamanu y Madre de Diós están aisladas de sus cauces principales, mientras que todas las lagunas del rÃo Manuripi están conectadas a su cauce principal.
Muestreo. Se consideraron dos categorÃas a escala espacial. La primera, la 'Cuenca' que es una escala espacial grande, en esta categorÃa se consideran a los rios: Tahuamanu, Manuripi y Madre de Dios. La segunda categorÃa es el 'Hábitat' que es una escala espacial pequeña, en esta categorÃa se consideran al RÃo (los cauces) y la Laguna (las aguas estacionarias en las zonas de inundación).
Las muestras fueron tomadas en cuatro sitios del RÃo y Laguna del Tahuamanu, tres sitios del RÃo y Laguna del Manuripi, además tres sitios del RÃo y dos sitios de la Laguna del Madre de Dios, sumando 19 sitios en el área del estudio, tal y como se muestra en la Figura 1. Todos los sitios fueron muestreados durante la estación seca, del 30 de mayo al 7 de julio de 2001.
Las variables fÃsico-quÃmicas registradas en la superficie de las aguas fueron: temperatura (C°), transparencia (Disco de SecchÃ, cm), pH, conductividad (uS cm-1) y oxÃgeno disuelto (mg L-1). En cada sitio, los datos fueron tomados después de la captura de peces entre 9:00 am y 10:00 am. Los datos del RÃo se obtuvieron en medio tramo, en donde se colocaron las redes agalleras; los datos de la Laguna se tomaron en el centro.
Las muestras de peces fueron colectadas con una baterÃa de 13 redes agalleras de rombos diferentes: 10, 15, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, y 110 mm de longitud entre nudo y nudo, 25 m de largo y 2.5 m de alto. En cada sitio se colocaron las redes durante dos horas de 5:30 a 7:30 am y dos horas de 5:30 a 7:30 pm aproximadamente. En el RÃo se colocaron las redes agalleras en las partes calmadas o remansos de orillas hacia las partes más profundas. En la Laguna, se colocaron las redes agalleras desde las orillas hacia las partes más profundas del centro, para garantizar la captura de mayor número de peces.
Los especÃmenes garantes fueron conservados en formol al 4% en el campo y después en alcohol etÃlico al 70% en el Museo IctÃcola del Centro de Investigación de Recursos Acuáticos (CIRA), Facultad de Ciencias Pecuarias, Universidad Autónoma del Beni "José Ballivián", Trinidad, Beni, Bolivia.
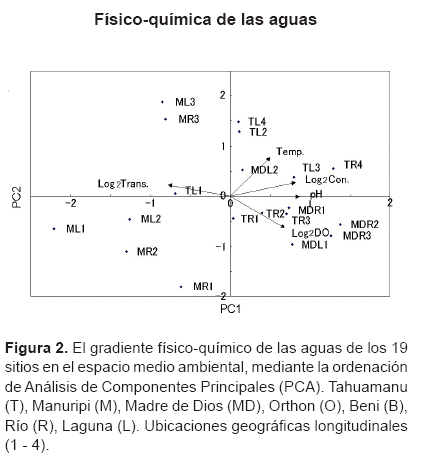
Análisis de los datos. Para identificar los mayores gradientes fÃsico-quÃmicos de aguas en el área de estudio se realizó el Análisis de Componentes Principales (PCA) con los datos fÃsico-quÃmicos de 19 sitios. Para ajustar la normalidad, los valores de oxÃgeno disuelto,transparencia y conductividad fueron transformados por el log2. Los valores de pH y temperatura no se transformaron, ya que ninguna transformación por el log2, ln ni log10 se ajustó la normalidad. Para compensar las diferentes unidades de registro, se extrajeron los componentes principales de la matriz de correlación y para determinar el número de ejes válidos a interpretar, se aplicó el análisis de 'broken stick eigenvalues'.
Para identificar los mayores gradientes de comunidades ictÃcolas en el área de estudio, se realizó el Análisis de Correspondencia Segmentado (DCA) con los datos de peces de 19 sitios. Los datos de entrada fueron la captura-por-unidad-esfuerzo (CPUE) en la abundancia numérica por especie de 19 sitios, datos transformados por el log2 (CPUE+1). La unidad de esfuerzo fue la combinación de las 13 redes agalleras colocadas durante dos horas en la mañana y dos horas en la tarde.
Para confirmar las relaciones entre los mayores gradientes fÃsico-quÃmicos de aguas y comunidades ictÃcolas en el área de estudio, se analizó la correlación entre los puntos de 19 sitios de PCAy los puntos de 19 sitios de DCA.
Las abundancias relativas de taxones superiores (abundancia medida como la proporción de CPUE, transformada por el arcoseno P0.5) entre las comunidades ictÃcolas discretas, fueron comparadas mediante el ANOVA con la prueba de Tukey en tres categorÃas de especies, distribuidas en uno, dos y tres hábitats. La distorsión entre diferentes categorÃas podrÃa considerarse como el balance de tácticas entre la adaptabilidad a condición óptica y la dispersión.
Como una prueba general del modelo PTM, se utilizarón las figuras de dispersión para ilustrar las relaciones entre la abundancia relativa de taxones superiores (transformada por el arcoseno P0.5) y la transparencia de aguas (transformado por el ln). La Regresión Localmente Ponderada (LOWESS) fue usada para resumir los patrones de dependencia entre la variable x y la variable y en las figuras de dispersión.
RESULTADOS
Gradiente fÃsico-quÃmico de las aguas. El primer y segundo Componente Principal (PC1 y PC2) del PCA, extrajeron 56,3% y 22,3% respectivamente de la variación fÃsica - quÃm ica de aguas en 19 sitios. El primer componente principal valió la pena ser interpretado como el gradiente fÃsico-quÃmico de aguas, considerando a la correlación limnológica muy conocida y los 'broken stick eigenvalues'. El PC1 también podrÃa ser interpretado como el gradiente de los cambios y las influencias de los diferentes tipos de aguas, ya que los puntos bajos de este eje fueron asociados con los valores bajos de conductividad y pH, y los valores altos de la transparencia. Por otro lado, los puntos altos de este eje fueron asociados con los valores altos de la conductividad y el pH, y los valores bajos de la transparencia (Tabla 1).
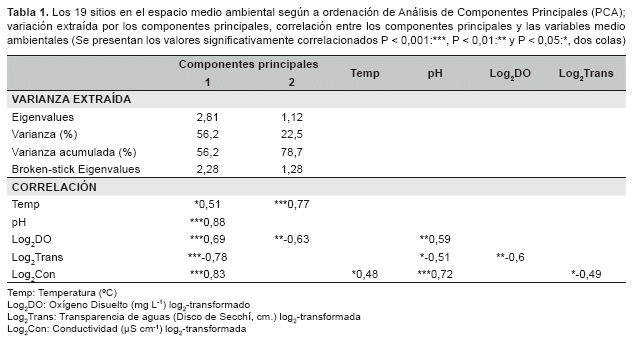
La dispersión de 19 sitios en el PC1 indicó que el gradiente de los cambios y la influencia de los diferentes tipos de aguas estaba cubierto entre los diversos hábitats del sistema fluvial y el posible patrón espacial geológico o vegetal de origen de valles en la escala espacial grande, asà como el mayor gradiente de los cambios y la influencia de los diferentes tipos de aguas, ya que los sitios de Manuripi tuvieron las aguas más negras que los de Tahuamanu y Madre de Dios. Por otra parte, el patrón espacial geomorfológico en la escala espacial pequeña o en los valles de aguas más blancas, Tahuamanu, Madre de Dios, asà como el menor gradiente de los cambios y las influencias de los diferentes tipos de aguas, ya que los sitios de Laguna tuvieron las aguas más negras que los de RÃo, Tahuamanu y Madre de Dios. Sin embargo, no indicó el patrón espacial geomorfológico en la escala espacial pequeña o en el valle de aguas más negras, Manuripi, ya que los sitios del RÃo y Laguna tuvieron las mismas aguas negras (Figura 2).
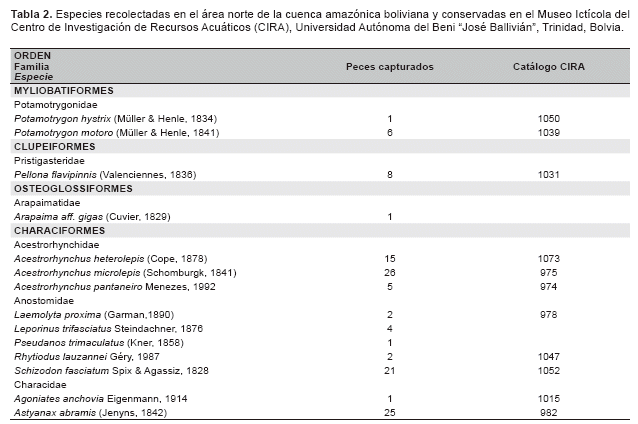
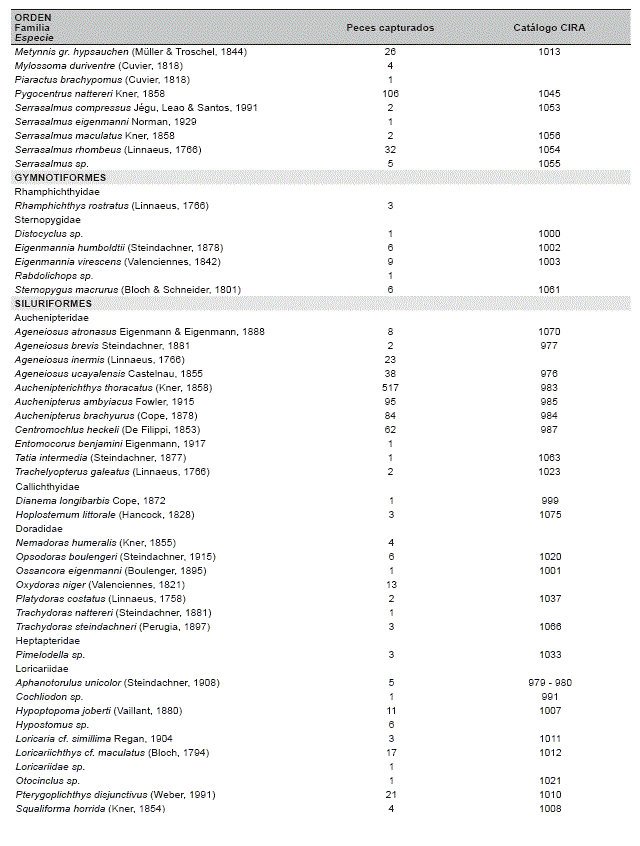
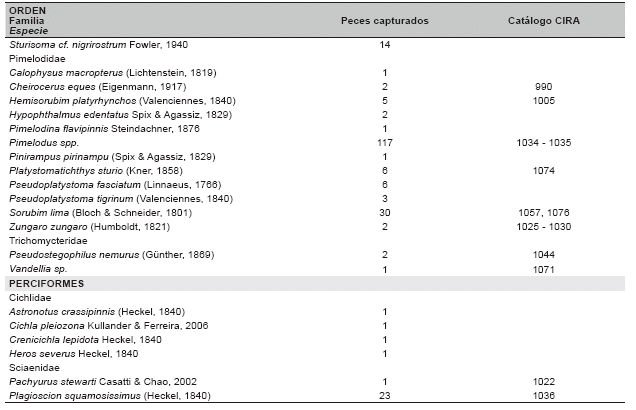
Gradiente de comunidades ictÃcolas. Se colectaron un total de 2 716 especÃmenes, pertenecientes a 129 especies, 24 familias y 7 órdenes, con un esfuerzo de 1 047 horas * 62.5 m2 (Tabla 2).

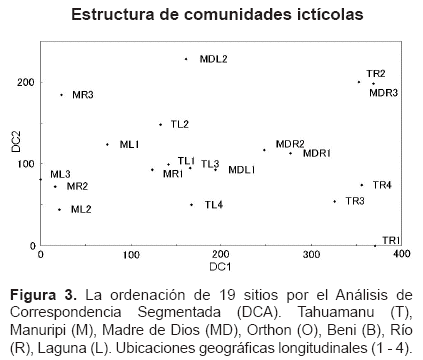
Como se muestra en la Figura 3, el primer eje de DCA (DC1) representa el mayor gradiente de comunidades ictÃcolas, considerando al largo de gradiente de 3,71 SD.
La dispersión de 19 sitios en el DC1 indica que el mayor gradiente de comunidades ictÃcolas estaba cubierto por los diversos hábitats del sistema fluvial, además el posible patrón espacial geológico o vegetal de origen de los valles en escala espacial grande como el mayor gradiente de comunidades ictÃcolas, ya que los sitios de Manuripi se dispersaron al lado izquierdo, y los sitios de Tahuamanu, Madre de Dios al derecho. Asà mismo, sugiere el patrón espacial geomorfológico en escala espacial pequeña como el menor gradiente de comunidades ictÃcolas, ya que los sitios de Laguna de Tahuamanu, Madre de Dios se dispersaron al lado izquierdo, y los sitios de RÃo de Tahuamanu, Madre de Dios al derecho. Sin embargo, no se observa el patrón espacial geomorfológico en escala espacial pequeña, ya que los sitios del RÃo y Laguna de Manuripi se sobrepusieron.
La dispersión de los sitios en el DC1 también indica la presencia de comunidades ictÃcolas discretas entre tres zonas: Manuripi, Laguna y RÃo de Tahuamanu, y Madre de Dios. Los Procedimientos de la Permutación Multi-Respondidos (MRPP) mostraron que la diferencia de comunidades ictÃcolas entre estas tres zonas fue significativa (p<0,001).
En la Figura 3 también se observa el segundo eje de DCA (DC2) que representa el menor gradiente de comunidades ictÃcolas, considerando al largo de gradiente de 2,28 SD.
Relación entre el gradiente fÃsico-quÃmico de las aguas y el gradiente de las comunidades ictÃcolas. El análisis de correlación entre los puntos de 19 sitios en PC1 y DC1 fue significativa (r=0,77; P=0,000; dos colas), mientras la correlación entre los puntos de 19 sitios en PC1 y DC2 no fue significativa (r=0,13; P=0,586; dos colas), indicando que el mayor cambio de comunidades ictÃcolas coincidió espacialmente con el cambio de tipos de aguas.
Los puntos en DC1, promedio (±SD), de 67 especies de los Characiformes 120 (±19,0), y 52 especies de los Siluriformes y los Gymnotiformes 183 (±27,2) fueron ligeramente diferentes (t= -2,17; P=0,060; ANOVA, Pruebas de Contraste, no asume las variaciones iguales, dos colas), asà mismo los puntos en DC1 de cuatro especies de los Cichlidae -13 (±24,5) y dos especies de los Sciaenidae 326 (±40,0) fueron significativamente diferentes (t= -7,22; P=0,024; ANOVA, Pruebas de Contraste, no asume las variaciones iguales, dos colas) (Figura 4).
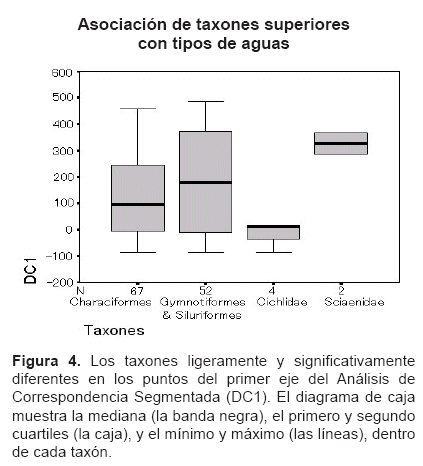
CaracterÃsticas de los hábitats. Los gradientes de aguas y peces, asà como su correlación permite identificar tres comunidades ictÃcolas discretas distribuidas en los tres hábitats de diferentes tipos de aguas: el RÃo de Tahuamanu y Madre de Dios, la Laguna de Tahuamanu y Madre de Dios, y el conjunto del RÃo y la Laguna de Manuripi, correspondiente a cauces de aguas blancas, lagunas de varzea, y valle de aguas negras, respectivamente (Tabla 2, Figuras 2 y 3).
Se colectaron 129 especies de peces, los mismos que fueron similares entre los valles de diferentes tipos de agua, 89 en aguas blancas y 85 en aguas negras. 44 especies fueron restringidas en valles de aguas blancas, 40 especies en valle de aguas negras y 45 especies distribuidas en ambos valles. La riqueza local (número de especies local) y la abundancia local (número de individuos local) fueron superiores en el valle de aguas negras que en los valles de aguas blancas, 28 (±8,1) vs 21 (±7,2) especies, 184 (±167,8) vs 113 (±89,6) individuos respectivamente. La riqueza y la abundancia local fueron superiores en las lagunas de varzea que en los cauces de aguas blancas, 27 (±5,0) vs 16 (±5,0) especies, 191 (±70,0) vs 46 (±25,4) individuos respectivamente, asà la riqueza y abundancia local fueron similares entre las lagunas de varzea y el valle de aguas negras. La alta riqueza y abundancia local del valle de aguas negras se debe a las especies de distribución restringida, por el contrario, en las lagunas de varzea se debe a las especies con amplia distribución (Tabla 3).
Las especies de distribución restringida son raras con bajas ocurrencias y abundancias. Las 56 especies fueron colectadas en los cauces de aguas blancas, de las cuales 15 fueron restringidas en este hábitat, más de una cuarta parte. El valor promedio de riquezas locales encontradas en este hábitat fue de 16 especies, de las cuales tres fueron restringidas, menos de una quinta parte. El valor promedio de abundancias locales encontradas en este hábitat fue de 46 individuos, de los cuales cuatro fueron de especies restringidas, que corresponde a menos de una decima parte. Este patrón fue común en los tres hábitats (Tabla 3).
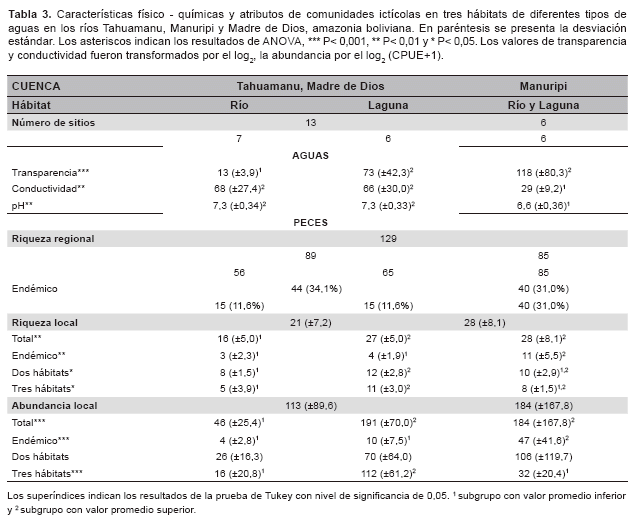
En términos de abundancia total, de 129 especies, 15 (1,1%) fueron colectadas solamente en cauces de aguas blancas, 15 (2,2%) en lagunas de varzea,40 (10,9%) en valle de aguas negras, mientras que 41 (47,9%) se encontraron en dos hábitats y 18 (37,9%) en tres hábitats.
El área de estudio presentó un stock regional de riqueza ictÃcola compuesta de 129 especies. En orden de importancia, 67 especies de Characiformes (51,9%), 46 especies de Siluriformes (35,7%), seis especies de Gymnotiformes (4,7%), cuatro especies de Cichlidae (3,1%) y dos especies de Sciaenidae (1,6%). Los taxones superiores que presentaron mayor riqueza poseen especies con amplia distribución, por ejemplo, solamente los Characiformes y los Siluriformes poseen especies recolectadas en los tres hábitats. Especies del orden Gymnotiformes no se recolectaron en los tres hábitats, aunque pueden estar distribuidas en estos hábitats. Los Cichlidae solo fueron colectados en valles de aguas negras y los Sciaenidae solo en valles de aguas blancas (Tabla 4).
Las proporciones de grupos taxonómicos superiores en las abundancias locales, de cierto modo, se ajustan al modelo PTM, los Characiformes y los Cichlidae, visualmente orientados, dominan en aguas claras, en lagunas de varzea y en valle de aguas negras. Por otro lado, los Gymnotiformes y los Siluriformes, adaptados a su baja visibilidad, dominan en aguas turbias y en cauces de aguas blancas. Los Sciaenidae están distribuidos principalmente en lagunas de varzea. Sin embargo, la tabla 4, también indica que la proporción de Characiformes y Siluriformes entre lagunas de varzea y valle de aguas negras presenta controversia al modelo PTM, dominando los Characiformes en lagunas de varzea con aguas más turbias y los Siluriformes en valle de aguas negras con aguas más claras. Las especies restringidas en su distribución presentan mayor ajuste al modelo PTM, los Characiformes dominan claramente en valle de aguas negras, y los Siluriformes en cauces de aguas blancas.
Los Gymnotiformes que carecen de especies con amplia distribución, están distri-buidos principalmente en valles de aguas blancas, asà mismo las dos especies de Sciaenidae están distribuidas solamente en valles de aguas blancas y las cuatro especies de Cichlidae están restringidas en valle de aguas negras. Por otro lado, las especies con mayor rango de distribución no se ajustan al modelo PTM; las especies de Characiformes y Siluriformes distribuidas en tres hábitats dominan en lagunas de varzea.
Tabla 4. Proporción de grupos taxonómicos superiores de las comunidades ictÃcolas en tres hábitats de diferentes tipos de aguas, rÃos Tahuamanu, Manuripi y Madre de Dios, amazonia boliviana. En paréntesis se presenta la desviación estándar. Los asteriscos indican los resultados de ANOVA, *** P < 0,001, ** P < 0,01 y * P < 0,05. Los valores de proporción fueron transformados por el arcoseno P0.5.

Los superÃndices indican los resultados de la prueba de Tukey, con nivel de significancia de 0,05. 1subgrupo con valor promedio inferior
y 2 subgrupo con valor promedio superior.
Luego, las especies de Characiformes distribuidas en dos hábitats dominan en valles de aguas blancas, y las especies de Siluriformes distribuidas en dos hábitats dominan en cauces de aguas blancas y valle de aguas negras.
También se tiene que, las especies tÃpicas restringidas en cauces de aguas blancas, en orden de importancia, según la abundancia total, fueron seis especies de Siluriformes: Platystomatichthys sturio con seis individuos, Aphnotorulus unicolor con cinco, Trachydoras steindachneri con tres, Pseudostegophilus nemurus con tres, Zungaro zungaro con dos y Cheirocerus eques con dos. De las nueve especies restantes solo se recolectó un individuo.
Las especies tÃpicas restringidas en lagunas de varzea fueron nueve, pertenecientes a diferentes taxones superiores: Pterygoplichthys disjunctivus con 20 individuos, Pellona flavipinnis con siete, Potamotrygon motoro con seis, Hypostomus sp. con seis, Leporinus trifasciatus con cuatro, Hoplosternum littorale con tres, Pseudoplatystoma tigrinum con tres, Hypophthalmus edentatus con dos, Serrasalmus maculatus con dos. De las seis especies restantes se colectó solo un individuo.
Las especies tÃpicas restringidas en valle de aguas negras fueron seis especies de Characiformes: Moenkhausia cf. lepidura I con 99 individuos, Moenkhausia cf. lepidura II con 26, Acestrorhynchus microlepis con 26, Metynnis gr. hypsauchen con 23, Acestrorhynchus heterolepis con 13 y Bryconops melanurus con 11, También se recolectaron 34 especies con menos de 10 individuos y 18 especies solamente con uno.
Las especies tÃpicas generalistas distribuidas en tres hábitats fueron 15, de las cuales 11 son Characiformes y cuatro Siluriformes, Potamorhina altamazonica con 197 individuos, Pimelodus spp. con 107, Psectrogaster rutiloides con 105, Pygocentrus nattereri con 96, Auchenipterus ambyiacus con 90, Triportheus angulatus con 80, Rhaphiodon vulpinus con 70, Anodus elongatus con 47, Prochilodus nigricans con 35, Sorubim lima con 29, Hydrolicus scomberoides con 25, Astyanax abramis con 24, Potamorhina latior con 21, Loricariichthys cf. maculatus con 16, Cynopotamus amazonus con 15. Tres especies presentaron menos de 10 pero más de cuatro individuos.
Las especies tÃpicas distribuidas en dos hábitats fueron 19, de las cuales 10 son Characiformes, ocho Siluriformes y una Sciaenidae: Auchenipterichthys thoracatus con 492 individuos, Moenkhausia dichroura con 142, Auchenipterus brachyurus con 78, Centromochlus heckeli con 62, Thoracocharax stellatus con 43, Roeboides affinis con 41, Ageneiosus ucayalensis con 38, Psectrogaster curviventris con 34, Serrasalmus rhombeus con 29, Steindachnerina leucisca con 23, Plagioscion squamosissimus con 22, Roeboides dispar con 22, Ctenobrycon hauxwellianus con 21, Schizodon fasciatum con 20, Ageneiosus inermis con 20, Curimatella meyeri con 14, Sturisoma cf. nigrirostrum con 13, Oxydoras niger con 13 e Hypoptopoma joberti con 10. Se colectaron 22 especies con menos de 10 individuos y cinco especies con solo dos individuos.
Prueba general del modelo PTM. Las figuras de dispersión y curvas de regresión LOWESS indican que los Characiformes fueron dominantes en aguas más claras, mientras que Siluriformes, Gymnotiformes y Sciaenidae fueron dominantes en aguas más turbias. Asà mismo, indican un umbral en x = 3 (20 cm de transparencia, Disco de SecchÃ) (Figura 5).
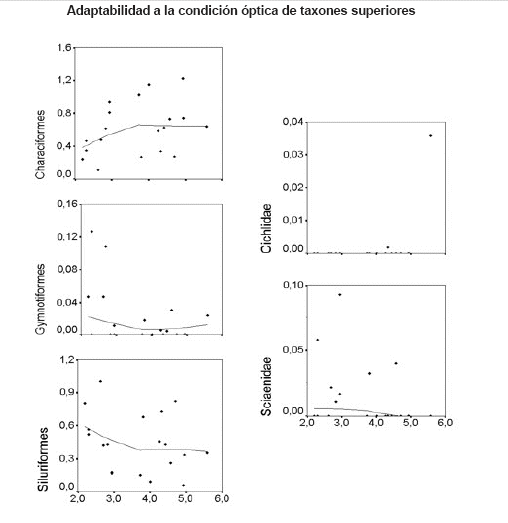
Figura 5. Abundancia relativa de taxones superiores (la abundancia medida como la proporción de CPUE, transformado por el arcoseno P0.5) respecto a la transparencia de aguas (Disco de SecchÃ, en cm transformado por el ln). Las lÃneas sólidas en los campos de datos son las curvas de regresión LOWESS.
DISCUSIÃN
Se identificaron tres comunidades ictÃcolas discretas, distribuidas en los hábitats con diferentes tipos de aguas: cauces de aguas blancas, lagunas de varzea y valle de aguas negras. La productividad por sà sola no es un indicador de la riqueza ni la abundancia de comunidades locales. Las lagunas de varzea y el valle de aguas negras presentan valores similares. Lo cual coincide con Henderson y Crampton (1997) quienes observaron que las lagunas de igapo podrÃan ser ambientes más estables que las lagunas de varzea, ya que la varzea tiene acumulación de sedimentos, asà la expansión y concentración de lagunas es mayor.
Por otro lado, las lagunas de igapo carecen de acumulación de sedimentos y están confinadas siempre en sus lechos, a pesar de la baja productividad las lagunas de igapo podrÃan albergar una alta densidad de peces. En el área de estudio se evidencia este fenómeno, casi todas las lagunas de varzea están aisladas mientras las lagunas de igapo están conectadas. Por otra parte, la alta riqueza y abundancia en las lagunas de varzea depende de especies con amplia distribución, mientras el valle de aguas negras depende de especies con distribución restringida.
Las especies restringidas en un hábitat muestran el mayor control filogenético, los Characiformes dominan en el valle de aguas negras y los Cichlidae están distribuidos solamente en este hábitat, mientras los Siluriformes dominan en los cauces de aguas blancas. Las especies distribuidas en tres hábitats muestran el menor control filogenético, tanto Characiformes como Siluriformes dominan en las lagunas de varzea. Las especies distribuidas en dos hábitats, algunos grupos taxonómicos superiores muestran el control filogenético y otros no. Los Gymnotiformes y los Sciaenidae están distribuidos en los valles de aguas blancas, sin embargo los Characiformes dominan en los valles de aguas blancas y los Siluriformes dominan en los cauces de aguas blancas y el valle de aguas negras. Los taxones superiores con mayor diversidad poseen las especies con mayor rango de distribución y menor control filogenético, las cuales son las especies dominantes a escala regional.
La filogenia y distribución de los Curimatidae sugieren el origen de mayorÃa de componentes previo a la división actual de principales cuencas hidrográficas (Vari, 1988, Vari & Weitzman, 1990). La filogenia y distribución de los Gymnotiformes destacan el control filogenético en tácticas para adaptarse a los diferentes hábitas más que la división actual de principales cuencas hidrográficas, reconociendo (i) pequeños rÃos en pie de monte andino y escudo precámbrico, normalmente entre 200 y 1000 msnm, (ii) pequeños rÃos de tierra firme sobre el nivel de inundación anual en la formación terciaria, (iii) rÃos profundos con inundación anual, y (iv) zona de inundación en la formación cuaternaria, destacando el rol de diferentes tipos de aguas (Crampton, 2011).
La fragmentación de continente Gondwana se inició desde finales del Cretáceo. La historia posteriori de tierras en la región neotropical se sintetiza como la persistencia permanente de masas precámbricas, la elevación de Cordillera Andina y la formación de tierras bajas (Lundberg et al., 1998). Las evidencias en fósiles muestran la extinción de especies arcaicas y la especiación de la mayorÃa de formas hoy reconocidas como géneros o taxones superiores (tribus, subfamilias) hasta el Paleógeno. Durante el Neógeno la especiación ha sido limitada en taxones inferiores (Lundberg, 1998). El marco de habitas similares a lo actual fue establecido probablemente hasta el Paleógeno, asà la diferencia de tácticas para adaptarse a diferentes hábitats tiende a evidenciarse entre géneros o taxones superiores, mientras especies pertenecientes a mismo género tienden a presentar tácticas similares (Lopez-Fernández y Albert, 2011). Durante el Neógeno, el complejo ciclo de conexión y desconexión de divisiones hidrográficas originaron oportunidades para la especiación y extinción por aislamiento, y la dispersión (Albert y Carvalho, 2011).
Para lograr comprender la evolución en forma completa, es necesario abarcar la historia de peces de la región neotropical en un periodo de más de 120 millones de años. Durante este periodo, ciertas tácticas podrÃan haberse asociado a la diversidad actual, asà algunos taxones superiores son muy diversos y otros muy conservadores. En forma general, los grupos con mayor diversidad actual tuvieron orÃgenes antiguos, ancestros probablemente de menor porte corporal con limitada capacidad de dispersión, asà mismo podrÃan haber experimentado más frecuentemente la innovación de tácticas. Por el contrario, los linajes con mayor porte corporal se extinguieron o fueron representados por pocas especies como el Paiche (Arapaima)y Pez Pulmonado (Lepidosiren). La capacidad de dispersión podrÃa haber sido influenciada por tácticas de ancestros como el porte corporal y la especialización ecológica en función a ambientes abióticos y bióticos (Albert et al., 2011).
Los resultados sugieren un proceso de coevolución a gran escala espacio temporal, como los ancestros con menor porte corporal, adaptación a condición óptica especializada con limitada capacidad de dispersión, asà se fueron innovando las tácticas para adaptarse a un ambiente con mayor fluctuación estacional, a los habitats como las lagunas de varzea, logrando finalmente mayor porte corporal, adaptación a la condición óptica generalizada y alta capacidad de dispersión.
El inventario de peces está en pleno proceso de desarrollo en las tierras bajas de la amazonia boliviana (Pouilly et al. 2010, Carvajal y Fernández 2011, Hablützel et al. 2013), sin embargo se conoce muy poco sobre la distribución de estructuras de comunidades ictÃcolas para llevar a cabo cualquier tipo de acción de aprovechamiento sostenible.
Se conoce que las tierras bajas de la AmazonÃa boliviana están circunscritas por dos formaciones geológicas: Andes y Precámbrica. Los rÃos originarios de Andes son de aguas blancas, como el Madre de Dios, Beni y Mamoré, siendo la zona de inundación del rÃo Mamoré (varzea), una de las más extensas y productivas del Mundo. El rÃo de aguas claras como el Itenez es originario de la formación Precámbrica. En la parte Norte se encuentran numerosos arroyos que drenan a tierras firmes y que afluyen a rÃos de aguas negras como el Manuripi y Yata. En la parte Este, los arroyos que drenan a los bosques, afluyen en rÃos de aguas negras como el rÃo San MartÃn, estos rÃos de aguas negras presentan la zona de inundación (iguapo) (Roche etal., 1992; Yunoki et al., 2010; Yunoki y Torres, 2011).
Los estudios recientes en filogenia y biogeografÃa en la región Neotropical sugieren el rol de la heterogeneidad de hábitats para el origen y mantenimiento de la biodiversidad regional, además del estudio ecológico desde el punto de vista de la teorÃa de metacomunidad (Crampton, 2011). Las tierras bajas de la amazonia boliviana es un sistema continuo, sin embargo abarcan hábitats heterogéneos y especies adaptadas en diferente grado, a cada hábitat dispersado entre ellas.
El evidente desajuste al modelo PTM de lagunas de planicie en el Mamoré Central podrÃa deberse al aislamiento de lagunas por barreras fÃsicas (Pouilly y RodrÃguez, 2003). Las lagunas septentrionales son peculiares, siendo ubicadas en el cauce antiguo del rÃo Beni (Wilkinson et al., 2006) y conectadas por el arroyo negro y el rÃo Yata. No se conoce la dinámica espacial entre los valles de aguas negras que están separados de los valles de aguas blancas, como de los rÃos San MartÃn, Yata, Manuripi, etc., donde la condición óptica y la depredación por ictiófagos, podrÃan formar una barrera biológica de peces.
Losanálisisdedatosexistentes sobre ladistribución de estructuras de comunidades ictÃcolas (Yunoki et al. 2010, Yunoki y Torres 2011) y el trabajo de inventario en áreas aún no exploradas basado en instrumentos geográficos como ecoregión, en caracterÃsticas geomorfológicas de clima, cobertura vegetal, suelos y usos, etc., junto con unidades hidrográficas menores, todo esto podrÃa complementar los conocimientos de la dinámica espacial. Este campo emergente es probablemente significativo no solo para el estudio de ecologÃa básica de peces, sino también para la conservación y manejo de múltiples especies.
Si se considera que existe la dinámica espacial, entonces una barrera fÃsica artificial como una planta hidroeléctrica, o por otro lado, la introducción de especies exóticas, podrÃa tener consecuencias regionales en la diversidad de peces nativos.
CONCLUSIONES
La filogenia, la biogeografÃa y la teorÃa de meta comunidad indican que las tierras bajas de la amazonÃa pueden ser consideradas como manchas de hábitats con diferentes tipos de aguas (varzea, tierra firme e igapo), separadas por rÃos. El modelo PTM presenta mayor ajuste para especies restringidas y menor ajuste para especies con amplia distribución, indicando un balance de tácticas entre la adaptación a condición óptica y la dispersión. Los taxones superiores con mayor diversidad poseen especies con mayor rango de distribución y menor control filogenético, las cuales son especies dominantes a escala regional. Estas especies, al disminuir su adaptación a la condición óptica y aumentar su dispersión, han adquirido posiblemente una ventaja comparativa en la cadena trófica. La aplicabilidad del modelo PTM desde la perspectiva de la teorÃa de metacomunidad, sugiere inherentemente que el efecto de los depredadores sobre las poblaciones de presas no solo causan la mortalidad directa, sino también se tiene el comportamiento contra-predador, al igual que historias vitales por las cuales las comunidades ictÃcolas de diferentes tipos de aguas pueden coexistir y coevolucionar. La teorÃa de metacomunidad es probablemente relevante, no solo para el estudio de ecologÃa básica de peces sino también para la conservación y manejo de múltiples especies en las tierras bajas de la Amazonia.
AGRADECIMIENTOS
Agradecemos a las siguientes instituciones y personas: Universidad Amazónica de Pando. Julio A. Rojas Guamán, Director de CIPA. ONGs. HERENCIA. Luis Fernando Reyes, Romer S., Miserendino Salazar y Julio Carrasco Oshinaga. IRD Francia y NABU Alemania. Armada Boliviana acantonada en la ciudad de Cobija que colaboraron en la parte logÃstica. Guardias de la "Reserva Nacional de Vida Silvestre Amazónica 'Manuripi Heat' y a los estudiantes de la carrera de Piscicultura del Universidad Amazónica de Pando. Un agradecimiento especial a nuestro colega Alfredo Parada Montalvan por el trabajo incansable y conocimiento de pescador. Los colegas y autoridades de la UAB y a todas las personas asentadas a lo largo de los rÃos: Tahuamanu, Manuripi y Madre de Dios por su apoyo moral.
REFERENCIAS BIBLIOGRÃFICAS
AlbertJ.S., Bart Jr.H.L. &Reis R.E., 2011. Species Richness and Cladal Diversity, pp 89-104, In: Historical Biogeographyof Neotropical Freshwater Fishes, (ed.) Albert J. S. & Reis R. E. [ Links ]
Albert J.S. & Carvalho T.P., 2001. Neogene Assembly of Modern Faunas, pp 119-136, In: Historical Biogeographyof Neotropical Freshwater Fishes, (ed.) Albert J. S. & Reis R. E. [ Links ]
Carvajal-Vellejos F.M. y Fernández A.J.Z., 2011. Diversidad y distribución de los peces de la AmazonÃa boliviana, pp 101-147. En: Van Damme P. A., F. M. Carvajal y J. Molina (ed.) Peces y Delfines de la AmazonÃa boliviana, Habitas, potencialidad y amenazas, Cochabamba: Editorial INIA [ Links ]
Crampton W. G. R., 2011. An Ecological Perspective on Diversity and Distributions, pp 165-189, In: Historical Biogeographyof Neotropical Freshwater Fishes, (ed.) Albert J. S. & Reis R. E. [ Links ]
Fittkau E., Irmler U., Junk W., Reiss F. & Schmidt G., 1975. Productivity, Biomass, and Population Dynamics in Amazonian Water Bodies, pp 289-311. In: Amazonian Aquatic Systems. Springer-Verlag New York Inc. [ Links ]
Hablütel P I., Yunoki T. & Torres L.V., 2013. Update on the checklist of fish species of the Bolivian Amazon, Check List 9 (20) [ Links ]
Henderson P.A. & Crampton W.G.R., 1997. A com-parison of fish diversity and abundance between nutrient-rich and nutrient-poor lakes in the Upper Amazon, Journal of Tropical Ecology, 13, pp 175-198 [ Links ]
Jackson D.A., Peres-Neto P.R. & Olden J.D., 2001. What controls who is where in freshwater fish communities: the roles of biotic, abiotic, and spatial factors? Can. J. Fish. Aquat. Sci. 58, pp 157-170 [ Links ]
LeiboldM.A.,HolyoakM.,MouquetN.,Amarasekare P., Chase J.M., Hoopes M.F., Holt R.D., Shurin J.B., Law R., Tilman D., Loreau M. & Gonzalez A., 2004. The metacommunity concept: a framework for multi-scale community ecology, Ecology Letters, 7, pp 601-613 [ Links ]
Lopez-Fernandez H. & Albert J.S., 2011. Paleogene Radiations, pp 105-117, In: Historical Biogeography of Neotropical Freshwater Fishes, (ed.) Albert J. S. & Reis R. E. [ Links ]
Lundberg J.G., 1998. The temporal context for the diversification of Neotropical fishes, pp 49-68, In Phylogeny and Clasificaron of Neotropical fishes, (ed.) Malabarba L.R., Reis R.E., Vari R.P., Lucena Z.M.S. & Lucena C.A.S.
Lundberg J.G., Marshall L.G. Guerrero B.H. Malabarba M.C.S.L. &WesselinghF., 1998. The stageof Neotropical fish diversification: Ahistoriy of tropical South American rivers, pp 13-48, In Phylogeny and Clasificaron of Neotropical fishes, (ed.) Malabarba L.R., Reis R.E., Vari R.P., Lucena Z.M.S. & Lucena C.A.S.
Pouilly, M.& RodriguezM.A., 2003. Determinismof fish assemblage structure in neotropical floodplain lakes: influence of internal and landscape lake conditions, pp 243-264 [ Links ]
Pouilly M., Jégu M., Camacho J.T., Quintanilla M.P., Miranda G.C., Zubieta J.P.Z. y Yunoki T., 2010. Lista actualizada y distribución de los peces en las tierras bajas de la AmazonÃa Boliviana, Rev. Bol. Ecol. y Cons. Amb. 28, pp 73-97 [ Links ]
RodrÃguez, M.A. & Lewis Jr. W.M., 1997. Structure of fish assemblages along environmental gradients in floodplain lakes of the Orinoco river, Ecological Monographs 67, pp 109-128 [ Links ]
Roche M.A., Fernández J.C., Aliaga R.A., PeñaM.J., Salas R.E. & Montaño V.J.L., 1992. Balance hÃdrico superficial de Bolivia, PHICAB, CONAPHI-BOLIVIA, ORSTOM, UNESCO, IHH, SENAMI. 29 pp. [ Links ]
Saint-Paul U., Zuanon J., Correa M.A.V., GarcÃa M. & Fabré N.N., 2000. Fish communities in central Amazonian white and black water floodplains, Environmental Biology of Fishes 57, pp 235-250 [ Links ]
Servicio Nacional de HidrografÃa Naval, 1998. HidrografÃa de Bolivia, La Paz. Bolivia [ Links ]
Vari, R.P., 1988. The curimatidae, a lowland neotropical fish family (Pisces: Characiformes); distribution, endemism, and phylogenetic biogeography. Proceedings of a workshop on Neotropical distribution patterns, pp 343-377 [ Links ]
Vari, R.P. and Weitzman S.H., 1990. A review of the phylogenetic biogeography of the freshwater
fishes of South America. Vertebrates in the tropics Peters, G. and R. Hutterer Eds: 381-393
Wilkinson M.J., Marshall L.G. & Lundberg J.G., 2006. River behavior on megafans and potential influences on diversificaron and distribution of aquatic organisms, Journal of South American Earth Sciences 21, pp 151 -172 [ Links ]
Yunoki T., Torres L., Ten S. y GarcÃa V., 2010. Aguas y peces del TCO Itonama y Parque departamental-Ãrea natural de manejo integrado Itenez, Beni - Bolivia, Medio Acuático 1, pp 4-16 [ Links ]
Yunoki T. y Torres L., 2011. Aguas y peces del Mamoré Central, Beni - Bolivia, Medio Acuático, 2, pp 4-25 [ Links ]