








Services on Demand
Article
 Article in pdf format
Article in pdf format Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
 Cited by SciELO
Cited by SciELO
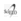 Similars in SciELO
Similars in SciELO
Bookmark
Revista Científica Agrociencias Amazonía
Print version ISSN 2307-9606
Rev.Cient.Agro.Amaz. vol.1 no.1 Beni May 2013
ARTICULO
Expresión del Gen Prosistemina de Tomate En Papa (Solanum tuberosum)
Araujo-Burgos T.1 y Rao R.2
1 Laboratorio de Biotecnología Vegetal, Carrera de Ingeniería Agronómica, Universidad Autónoma del Beni, Trinidad, Beni. Correo de contacto: agrotania@gmail.com
2 Departamento de Ciencias del Suelo, de la Planta, del Ambiente y de la Producción Animal, Universidad de los Estudios de Nápoles Federico II, Nápoles, Italia
RESUMEN
La sistem ina es una molécula de señal presente en el tomate y deriva de la región terminal C del precursor prosistemina. Involucrada en la respuesta local y sistémica de la planta a la masticación, regula la activación de diversos genes de defensa en respuesta al ataque de herbívoros en plantas de tomate. Homólogos de ésta molécula se han encontrado en otras Solanáceas como la papa, pero con menor actividad que la del tomate. Para conocer el rol biotecnológico de la prosistemina de tomate en la mejora de la resistencia endógena de la planta a situaciones de estrés biótico, el ADN complementario (ADNc) de prosistemina de tomate se expresó en papa, al igual que una prosistemina incompleta sin el exón que codifica para la sistem ina (Sys) para definir la posible función de la región terminal N del precursor en la activación de la respuesta de defensa. La transformación genética de papa se realizó mediante Agrobacterium y luego de la caracterización molecular de los transgénicos, se analizó un grupo de genes involucrados en la respuesta de la planta a estrés biótico mediante PCR en tiempo real. El análisis de expresión génica mostró que la modificación de la expresión de algunos genes en papa no se debe solo a la secuencia Sys, sino también a la terminal N de la prosistemina que jugaría un rol en la activación de algunos genes de defensa.
Palabras clave: sistemina, genes de defensa, papa transgénica, expresión génica
ABSTRACT
Systemin, a peptide isolated from tomato, derives from the C-terminal region of the prosystemin precursor. It is a signaling molecule involved in local and systemicwound response. It regulates the activation of several defensive genes in tomato plants in response toherbivoreattacks. Prosystemin homologues have been found in other Solanaceae species such as potato, but they were less active than the tomato one. The aim of this project was to increase the understanding of the possible biotechnologicalroleofthetomatoprosystemin, with a particular emphasis to the exploitation of this precursor to increase the endogenous resistance against biotic stress. To this goal, the tomato prosystemin ADNc was expressed in potato. Moreover, a mutated prosystemin ADNc lacking the systemin encoding exon (Sys) was also expressed to understand the possible function of the N-terminal region of the precursor in the activation of the defense response. Transgenic plants were obtained after an Agrobacterium mediated plant transformation. Potato transformants were characterized and a group of genes involved in plant response against biotic stress was analyzed by real time PCR. The gene expression analysis suggested that modification in gene expression in potato is not only due to the Sys sequence and that the N-terminal is also involved.
Key words: systemin, defense genes, transgenic potato, gene expression
INTRODUCCIÓN
Los mecanismos de defensa de la planta pueden ser de tipo constitutivo, es decir, que están siempre presentes en la planta, pero como esto incluye un costo metabólico no se puede mantener continuamente en el tiempo, así la planta ha desarrollado respuestas de defensa inducidas por la actividad alimentaria de los insectos como la síntesis de nuevos compuestos que obstaculizan los procesos digestivos y de absorción de nutrientes (Felton, 2005). Esta variedad de compuestos como inhibidores de proteasas, lipoxigenasas, compuestos orgánicos volátiles (VOC), terpenos, taninos, etc. comprometen el crecimiento del insecto fitófago (Ryan, 2000; Chen et al., 2005). Al respecto un papel central lo ocupa la vía metabólica del los octadecanoides, bien caracterizada en Solanáceas, la cual se induce mediante ataque de insectos fitófagos y termina produciendo especialmente ácido jasmónico y sus derivados (Walling, 2000). En el tomate una molécula señal de ésta vía es la sistemina, proteína de 18 aminoácidos que es liberada del precursor prosistemina (Pearce et al., 1991). El gen prosistemina contiene 11 exones, de los cuales el último codifica para sistemina y está ubicado en la región terminal C del precursor (McGurl et al., 1992). La molécula está involucrada en la respuesta local y sistémica de la planta a la masticación, además regula la activación más de 20 genes de defensa en respuesta al ataque de herbívoros en tomate (Ryan y Pearce, 2003). Igualmente, otros péptidos fueron aislados dentro de la familia Solanácea (Pearce y Ryan, 2003; Pearce et al., 2007; Pearce et al., 2009) con función similar a la prosistemina, glicopéptidos o sisteminas ricos en hidroxiprolina que inducen genes de defensa después del ataque de herbívoros (Narváez-Vásquez et al., 2007).
El presente trabajo se basa en el estudio de la expresión del gen prosistemina de tomate en papa y del mismo gen al cual se le ha borrado la porción que codifica para la sistemina. Los genotipos transgénicos de papa PR presentan la inserción del ADNc de prosistemina de tomate y los genotipos transgénicos PRx contienen el mismo gen incompleto. El objetivo de la investigación es ampliar el conocimiento sobre el rol biotecnológico de la prosistemina, sobretodo, la función de este precursor para incrementar la resistencia endógena de la planta frente al estrés biótico en una especie diferente al tomate.
MATERIALES Y MÉTODOS
Transformación genética de Agrobac-terium. Los vectores binarios PR (ADNc de prosistemina) y PRx (prosistemina incompleta) se encontraban disponibles en células de Escherichia coli cepa DH5a. La bacteria se inoculó en medio LB (Sambrook y Russell, 2001) con la adición de 15 g/l de agar bacteriológico y 50 ug/ml tetraciclina, las placas se incubaron por 16 horas a 37°C. Una sola colonia de bacteria transformada se inoculó en tubos de 100 ml con caldo LB y 50 ug/ml tetraciclina, luego se incubaron durante 16 horas a 37°C y 200 rpm de rotación. El ADN del plasmidio se aisló con el kit NucleoBond Midi Plasmid DNA purification (Macherey-Nagel). La integridad y la cantidad del ADN se controlaron mediante electroforesis en gel de agarosa al 1% con bromuro de etidio, en solución tampón TAE al 1%. El ADN de plasmidio conservado a -20°C se introdujo en Agrobacterium tumefaciens cepa LBA4404.
Las células competentes se obtuvieron por método químico. La bacteria se inoculó en caldo LB e incubó por 20 horas a 28°C y 200 rpm, luego 2 ml de ésta suspensión se adicionó a 50 ml de caldo YEP (10 g peptona, 10 g extracto de levadura, 5 g NaCl para 1 litro). El cultivo se colocó en cámara de crecimiento a 28°C y 200 rpm por ~8 horas hasta que la suspensión alcanzó una OD600~1.0. Luego se centrifugaron las células a 3000 rpm, a 4°C durante 5 min y se resuspendieron en 1 ml CaCl220 mM. Las células competentes obtenidas se dividieron en alícuotas de 1 ml y se conservaron a -80°C. Para la transformación, 1 ug de ADN plasmídico se añadió a cada uno de los tubos, las células se congelaron en nitrógeno líquido e inmediatamente se incubaron en baño María a 37°C por 5 min. Luego se añadió caldo LB a cada tubo y se incubaron a 28°C y 200 rpm por 2 horas. Las células se colectaron por centrifugación a 3000 rpm y el pellet se resuspendió en 0.2 ml de caldo LB. Las posibles células transformadas se inocularon en LB agar con los antibióticos selectivos (tetraciclina 5 ug/ml y rifampicina 50 ug/ml) e incubaron a 28°C por 3 días.
Control de células de Agrobacterium transgénico. Una colonia solitaria de las células cultivadas en LB agar con antibióticos se inoculó en 10 ml de caldo LB con los mismos antibióticos selectivos. Los tubos se incubaron a 28°C y 200 rpm por 24 horas. De cada tubo, 2 ml se conservaron en glicerol (300 ul de suspensión y 600 ul de glicerol al 50%) y congelaron a -80°C. Para controlar el transgen, se extrajo ADN plasmídico mediante protocolo mini-prep, se centrifugaron 8 ml de la suspensión a 4300 rpm por 10 min, el pellet de células se resuspendió en 533 ul de Solución I (glucosa 50 mM, TrisCl 25 mM, EDTA 10 mM, pH 8.0). Los tubos se enfriaron por 10 min y se añadieron 1100 Mi de Solución II (NaOH 0.2M y SDS 1%), después de mezclar y enfriar por 5 min, se añadieron 800 ul de Solución III (60 ml acetato de potasio 5M, 11.5 ml acido acético glacial y 28.5 ml agua bi-destilada), se enfrió y centrifugó. Todos los pasos de centrifugación se realizaron a 4°C, con 13200 rpm durante 5 min. Al sobrenadante se añadieron 500 ul de fenol-cloroformo (1:1), se centrifugó y al nuevo sobrenadante se le añadieron 10 ul ARNsa (10 mg/ml) lo cual se incubó a 37°C por 30 min. Luego se mezcló con 500 ul de fenol-cloroformo y al sobrenadante obtenido por centrifugación, se le añadieron 500 ul de cloroformo para centrifugar nuevamente. Al nuevo sobrenadante se le añadió 1 ml de etanol al 100% y luego se enfrío por 5 min. Después de centrifugar, el pellet de ADN colectado se lavó con etanol al 70%, se lo recuperó por centrifugación para finalmente resuspenderlo en 50 ul de agua bidestilada. La calidad y cantidad del ADN plasmídico se controló mediante electroforesis en gel de agarosa al 1% con bromuro de etidio, en tampón TAE al 1%. La solución de ADN se almacenó a -20°C para ser analizado por PCR. Luego se realizó el test de ketolactosa, basado en la diferencia metabólica entre el género Agrobacterium y Escherichia, para controlar la pureza de las colonias transformadas. Transformantes de A. tumefaciens y E. coli (control negativo) se inocularon en agar lactosa (lactosa 1%, extracto de levadura 0.1 % y bacto agar 2%) y se incubaron a 30°C por 3 días. Todas las colonias se cubrieron con solución de Benedict (132 g citrato de sodio, 256 g carbonato anhidro de sodio y 13.5 g de sulfato cúprico para 1 litro) e incubaron a temperatura ambiente por 30 min.
Transformación genética de papa. Solanum tuberosum, variedad Desirée creció in vitro en caldo MS20 (4.3 g mezcla de sales MS con vitaminas y 20 g sacarosa para 1 litro, pH 5.8) a 21 °C, fotoperiodo de 16 horas, 100 rpm, durante 4 semanas. Las cepas de Agrobacterium conservadas a -80°C se inocularon en LB agar con los mismos antibióticos selectivos, e incubaron a 28°C por 2 días. Una simple colonia se inoculó en caldo LB con antibióticos, las células crecieron a 28°C con ~200 rpm de rotación por 24 horas hasta que la concentración del cultivo alcanzó una OD600 entre 0.8 y 1.0. La virulencia de la bacteria se indujo usando el medio simplificado de inducción, SIM (citrato de sodio 20 mM, acetosiringona 0.3 mM y sacarosa 2%). El cultivo se centrifugó a 3000 rpm, el pellet se resuspendió en SIM e incubó a 28°C por 16 horas con movimiento rotacional; después se centrifugó a 3000 rpm y el pellet se resuspendió en 20 ml de caldo LB con los antibióticos selectivos para realizar el co-cultivo.
Las plantas de papa se transformaron como describen Narváez-Vásquez y Ryan (2002) con algunas modificaciones. Se cortaron entrenudos de 0.5 cm provenientes del tallo. Se colocaron los entrenudos sobre papel filtro que cubría el medio MS30 agar (4.3 g mezcla de sales MS con vitaminas, 30 g sacarosa, 8 g micro agar para 1 litro, pH 5.8) e incubaron a 22°C, fotoperiodo de 16 horas y en semioscuridad por 24 horas. A continuación, los entrenudos se sumergieron en la suspensión de células de Agrobacterium por 10 min y se colocaron en contacto directo con el medio MS30 a 22°C, 16 horas de fotoperiodo por 3 días en semioscuridad. Después se transfirieron a medio fresco P55 (4.3 g mezcla de sales MS con vitaminas, 30 g sacarosa, 8 g micro agar, 3 mg/l zeatina riboside, 3 mg/l acido giberélico y 0,1 mg/l acido indolacético para 1 litro, pH 5.8), con la adición de 200 mg/l cefotaxime para inhibir el crecimiento de Agrobacterium y 100 mg/l kanamicina para seleccionar transformantes. Las placas se mantuvieron a semisombra, 22°C y fotoperiodo de 16 horas. Apenas aparecieron los primeros brotes, se eliminó la semisombra y cuando éstos alcanzaron ~2 cm de largo se extrajeron para colocarlos en MS30 agar con 100 mg/l kanamicina hasta la emisión de raíces. Posibles transformantes que desarrollaron raíces en el medio selectivo, se propagaron in vitro y crecieron en MS30 agar con 50 mg/l kanamicina, a 22°C y 16 horas de fotoperiodo. Luego las vitroplantas se transfirieron a suelo estéril y crecieron bajo las mismas condiciones hasta la producción de tubérculos.
Material vegetal. Plantas transformadas y no transformadas de papa (Solanum tuberosum) variedad Desirée, crecieron a partir de tubérculos en invernadero de 23°C a 25°C y 16 horas de fotoperiodo. Plantas de 4 a 6 semanas de edad se utilizaron en todos los experimentos. Las líneas transgénicas debidamente caracterizadas, se seleccionaron a partir de la homogeneidad de los tubérculos.
Extracción y análisis de ADN genómico.
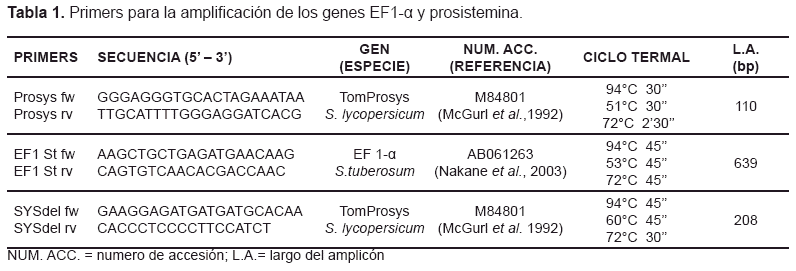
Se colectó material vegetal en nitrógeno líquido y se molieron 100 mg de hojas, el ADN genómico se aisló usando el kit Gen Elute Plant Genomic DNA miniprep (Sigma-Aldrich), luego se cuantificó mediante electroforesis en gel de agarosa al 1% con bromuro de etidio, comparando con un ADN Lambda (Promega) de cantidad conocida. Para la amplificación mediante Reacción en Cadena de la Polimerasa (PCR), 50 ul de solución madre se prepararon con 25-100 ng ADN, primers 0.4 uM, MgCl21.5 mM, dNTPs 100 uM y 1 U Taq ADN Polimerasa (Promega) en tampón PCR 1X (Promega). Los primers específicos reportados en la tabla 1 se usaron para la amplificación del gen prosistemina en los transformantes. La reacción de PCR se llevó a cabo en una Mastercycler Gradient (Eppendorf) con el ciclo termal correspondiente (tabla 1). Todos los productos de la amplificación mediante PCR y RT-PCR se observaron en gel de agarosa al 2% matizado con bromuro de etidio y se visualizaron con luz UV. Para la secuenciación de ADN, los productos de la amplificación (50 ul) se cargaron en gel de agarosa al 0.8% con el marcador 1Kb plus ADN Ladder (Invitrogen) para ver el tamaño del amplicón. Las bandas objetivo se cortaron hasta obtener 0,4 g de gel, después el ADN se aisló del gel con el kit QIAquick Gel Extraction (QIAGEN) para obtener 50 ul de solución de ADN y después de cuantificarlo, el ADN y los primers fueron enviados a la concentración y cantidad requerida por el servicio de secuenciación de ADN PRIMM (Nápoles). La secuenciación se hizo en el ADN Analyzer (ABI) Sequencer 3730 y las secuencias se analizaron con ayuda del software Clustal W2 (http://www.ebi.ac.uk/Tools/clustalw2/).
Extracción y análisis de ARN total. El ARN total se aisló de las hojas usando un método estándar de extracción en base a fenol/cloroformo. Las hojas colectadas se congelaron en nitrógeno liquido, 0.5 g de hojas se molieron y colocaron en tubos con 750 ul de tampón de extracción (Tris-HCl100 mM pH 8.5, NaCl100 mM, EDTA20 mM, SDS 1 %) y 750 ul de fenol-cloroformo (1:1). Todos los pasos de centrifugación se realizaron a 14000 rpm por 10 min a 4°C. Después de mezclar y centrifugar, la fase líquida se colectó y se repitió la extracción fenol/ cloroformo. El sobrenadante se transfirió a un tubo con 750 ul de cloroformo y se centrifugó; al nuevo sobrenadante se le añadieron 750 ul de isopropanol para que precipite el ARN. El pellet colectado por centrifugación se resuspendió en 200 ul de agua bidestilada para lavarlo, se precipitó mediante la adición de 1 volumen de LiCl 4M y enfrío por al menos 4 horas. Se centrifugó nuevamente por 15 min, el pellet de ARN se resuspendió en 400 ul de agua y se añadieron 40 ul de NaAc 3M pH 7.0 y 1 ml de etanol 96%. La solución se congeló y finalmente el pellet se recuperó por centrifugación. El ARN total se resuspendió por última vez en 42 ul de agua y cuantificó en espectrofotómetro, 2 ul de ARN total se diluyeron 1:100 y su concentración se midió a 260 nm; la pureza de ARN se verificó según la relación A260/A280.
La calidad de ARN se observó mediante electroforesis en gel de agarosa 1.2% en tampón desnaturalizante (400 ul formamida, 120 |jl formaldehido, 80 ul TAE 1%, 1.2 ul bromuro de etidio y 20 ul Loading Dye 6X); 4 ug de ARN en solución tampón se desnaturalizó a 65°C por 2 min antes de la electroforesis. Comprobada la óptima calidad del ARN, se adicionó ADNasa para eliminar cualquier contaminación de ADN, 10 ug de ARN total se trataron con 1U ADNasa/3ug ARN, (RNase-free DNase I, BioLabs), según las instrucciones del producto. El ARN se precipitó con la adición de 1/10 volúmenes NaAc 3M pH 7.0, 2 volúmenes de etanol 100%, y luego de enfriarlo, se centrifugó a frío por 15 min a 14000 rpm. El pellet se lavó con 1 volumen de etanol 70% y luego de enfriarse, se centrifugó a 14000 rpm por 10 min. El pellet resuspendido en 20 ul de agua, se lo usó para la transcripción inversa de ARNm con el kit RevertAid First Strand cDNA synthesis (Fermentas), siguiendo las instrucciones. El primer filamento de ADNc se sintetizó partiendo de 2 ug de ARN total. La calidad de la síntesis de ADNc se verificó usando los primers Elongation Factor-1 alpha (EF-1a) para S. tuberosum, diseñados para anillarse entre dos exones consecutivos y así detectar cualquier contaminación de ADN. Los primers específicos reportados en la tabla 1 se usaron para el control de la síntesis de ADNc y para amplificar el gen prosistemina en orden de verificar la transcripción de los transgenes. La reacción de PCR se preparó en 25 ul con 1 ul ADNc patrón, primers 0.4 uM, MgCl21.5 mM, dNTPs 100 uM y 0.5 U Taq DNA Polymerase (Promega) en tampón para PCR 1X (Promega). El ciclo térmico fue el que se indica en la tabla 1, realizado en una Mastercycler Gradient (Eppendorf), con 30 ciclos de amplificación. Los productos de amplificación se controlaron mediante electroforesis en gel de agarosa al 2%.
Extracción y análisis de proteínas totales.
0.5 g de hojas de papa congeladas en nitrógeno líquido se molieron hasta obtener un polvo fino. Un litro de solución de tampón salino fosfato (PBS) 10X se preparó con 87 g NaCl, 22.5 g Na2HPO4 y 2 g KH2PO4, pH 7.4 (Sambrook y Russell, 2001). Las proteínas se extrajeron en 300 ul de tampón PBS diluido 1:10, y conteniendo NaCl 0.5 M, EDTA 10 mM, PMSF 1 mM. La suspensión se mezcló vigorosamente en vortex y las muestras se centrifugaron a 13000 rpm, a 4°C por 20 min. El sobrenadante compuesto por proteínas solubles totales se cuantificó mediante espectrofotometría con el método Bradford y albumina de suero bovino (BSA) como proteína estándar (Bradford, 1976).
Para la electroforesis en gel de poliacrilamida con dodecilsulfato sódico (SDS-PAGE) las proteínas solubles totales se colocaron en geles de apilamiento al 5%, preparados con 670 ul de acrilamida 30%, 500 ul de Tris 1.0 M pH 6.8, 40 |jl de SDS 10%, 40 ul de persulfato de amonio 10% y 4 ul de TEMED en 4 ml. Los geles de resolución al 12% se prepararon con 8 ml de acrilamida 30%, 5 ml de Tris 1.5 M pH 8.8, 0.2 de SDS 10%, 0.2 de persulfato de amonio 10%, 8 ul de TEMED y agua hasta 20 ml. Un volumen de 15 ul conteniendo 40 ug de proteínas solubles totales se mezcló con igual volumen de tampón de muestra 2X (0.8 ml de tris-HCl 0.5 M pH 6.8, 2 ml glicerol, 0.2 ml de bromofenol azul 0.01% y volumen final de 10 ml). Estas muestras se desnaturalizaron a 100°C por 5 min antes de ponerlas al gel. Se usó la solución tampón de carrera Tris-Glicina-SDS 5X (94 g Glicina, 15.1 g Tris, 50 ml SDS 10% para 1 litro, pH 8.8) diluida 1X. La carrera electroforética se realizó en la Mini Protean Tetra Cell (BioRad) a 100 voltios por ~1 hora. Con el Precision Plus Protein standard (BioRad) se determinaron los pesos moleculares. Así, se obtuvieron dos geles idénticos para el análisis Western, uno se tiñó con azul brillante de Coomassie para visualizar proteínas, el otro gel se usó en la transferencia electroforética con membranas de nitrocelulosa (BioRad) para el Western blot.
La transferencia de las proteínas se llevó a cabo en el Trans-Blot Tranfer Medium (BioRad) con el tampón de transferencia 1X (Trizma 50 mM, Glicina 380 mM y metanol 10%) a 100 voltios por 1 hora. En seguida, se bloquearon los sitios no específicos sumergiendo la membrana de nitrocelulosa en una solución tampón A (PBS 1X con leche en polvo descremada 5% y Tween20 0.5%) por 1 hora. Todos los pasos de incubación y lavado se hicieron con movimiento de rotación suave, a temperatura ambiente. Luego, las membranas se incubaron por 3 horas con el anticuerpo primario anti-prosistemina (Delano etal., 1999) diluido 1:1000 en solución tampón Ay se lavaron en PBS 1XconTween20 0.5%. Después se incubó por 1 hora con el anticuerpo secundario, peroxidasa etiquetada con anti-IgG de conejo (Amersham), diluído 1:10000 en solución tampón A, se lavaron de igual modo adicionando un último lavado con PBS 1X por 10 min. Las proteínas de prosistemina se observaron con el kit ECL Western Blotting Detection System (Amersham) a 30 min de exposición, siguiendo las instrucciones del kit.
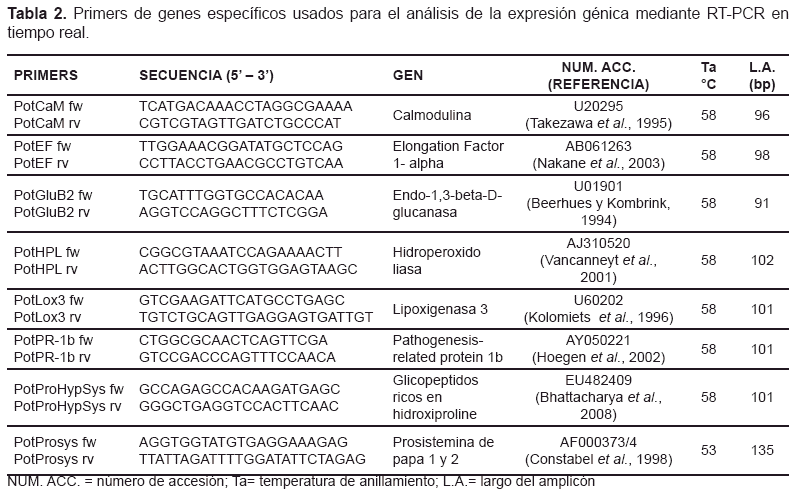
Análisis de expresión génica. Se llevó a cabo mediante RT-PCR en tiempo real, en el Fast Real-Time PCR System 7900 (Applied Biosystem) con 12,5 ul de Power Sybr Green PCR Master Kit 2X (Applied Biosystem), 0,3 ul del primer filamento de ADNc, 0,35 uM de primers específicos y agua hasta un volumen de25ul.
El ciclo termal comenzó con 50°C por 2 min, 95°C por 4 min, 40 ciclos de 95°C por 15 seg y 1 min a la temperatura de anillamiento de los primers (tabla 1). Se analizaron 3 réplicas biológicas por cada genotipo y 3 repeticiones para cada evento de amplificación. Para el control de los productos de amplificación se incluyó un paso de disociación con 15 seg a 95°C, 15 seg a 58°C y una desnaturalización lenta a 95°C por 15 seg, para comparar las curvas obtenidas con la Tm de cada par de primers mostrados en la tabla 2. Los productos también se observaron en gel de agarosa al 2%, cuyo tamaño de amplicón debe ser igual a los indicados en la tabla 2. Los datos emitidos fueron analizados mediante el software 7900HT Sequence Detection Systems, versión 2.3 (Applied Biosystem), usando el método comparativo de Ct con la formula ![]() (Livaky Schmittgen, 2001), donde:
(Livaky Schmittgen, 2001), donde:
![]()
El gen Elongation Factor 1-alpha (Nicot et al., 2005) fue el gen endógeno de referencia usado para normalizar los niveles de expresión de los genes objetivo en papa. Los primers de los genes específicos reportados en la tabla 2 se usaron para el análisis de la expresión génica de las plantas transgénicas. Estos se diseñaron con el software Primer Express 2.0 (Applied Biosystems) y el fragmento amplificado se eligió entre 90 y 135 pares de bases (bp).
Análisis estadístico. Los datos recolectados se analizaron mediante one-way ANOVA, seguido por Tukey HSD post-hoc-test para comparaciones múltiples. Todos los análisis se hicieron con ayuda del software SPSS, versión 16.0
RESULTADOS Y DISCUSIÓN
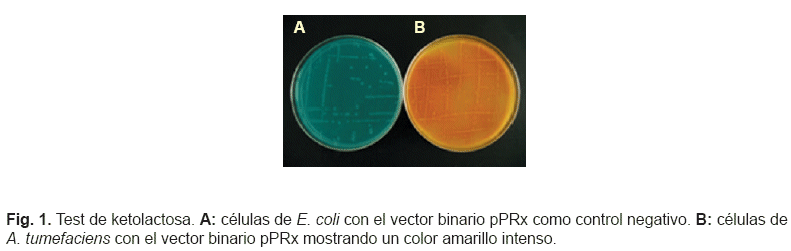
Transformación vegetal y caracterización molecular de los transgénicos. Los vectores binarios pPR con el casete de expresión que contenía el ADNc de prosistemina de tomate bajo el control del promotor CaMV35S, y el vector binario similar pPRx con el ADNc de prosistemina sin el exón que codifica para la sistemina, se insertaron en la bacteria A. tumefaciens, cepa LBA4404. Las posibles colonias transformadas que crecieron en medio selectivo se controlaron mediante PCR (no mostrado) y mediante el test de ketolactosa o de Benedict (fig. 1) donde se observó un anillo amarillo de Cu2O en colonias puras deAgrobacterium debido a la producción de 3-ketolactosa.
Luego, las bacterias con ambos vectores binarios se usaron para la transformación de Solanum tuberosum, variedad Desirée. Un ejemplo de entrenudos 7 semanas después del co-cultivo se muestra en la fig. 2. Luego de la formación de los callos, los brotes diferenciados se cortaron y transfirieron al medio selectivo hasta el enraizamiento. Los transformantes se nombraron como los constructos, PR y PRx.

Posibles transformantes que fueron capaces de desarrollar raíces en medio selectivo se analizaron mediante PCR para detectar la presencia del transgen de prosistemina de tomate. Los primers usados fueron Prosys para distinguir el ADNc completo de prosistemina y Sysdel para amplificar la prosistemina incompleta. La electroforesis en gel de agarosa de la PCR de los transformantes se muestra en las figuras 3 y 4.
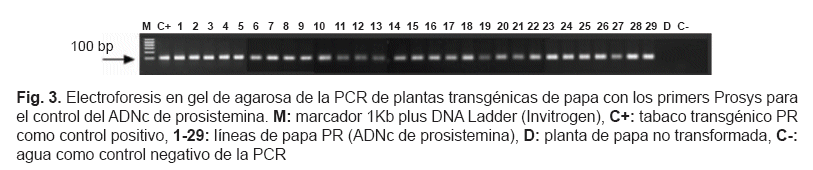

Un total de 29 líneas PR y 29 líneas PRx fueron positivas en el análisis PCR, todos los transformantes positivos se transfirieron in vivo en invernadero. A las 4 semanas, el ARN total se extrajo y la síntesis de ADNc se controló usando los primers Elongation Factor-1 alpha, diseñados para anillar en dos exones consecutivos y detectar una posible contaminación de ADN genómico. La transcripción de ambos transgenes fue controlada usando los primers Prosys y Sysdel mediante RT-PCR. La electroforesis del gel de agarosa de la RT-PCR de los transformantes se muestra en la fig. 5 y 6.

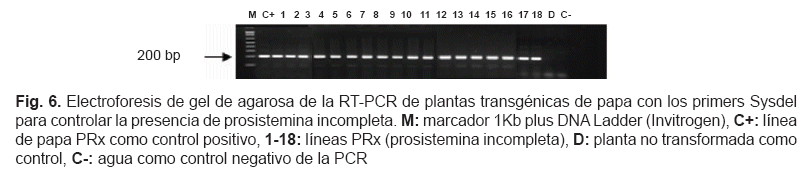
En 13 líneas PR, el transgen prosistemina se transcribía y 18 líneas PRx fueron positivas al mismo análisis. De todos los transformantes RT-PCR positivos se obtuvieron tubérculos para análisis posteriores. Los datos de la transformación genética de papa incluyendo el análisis molecular se muestran en la tabla 3.

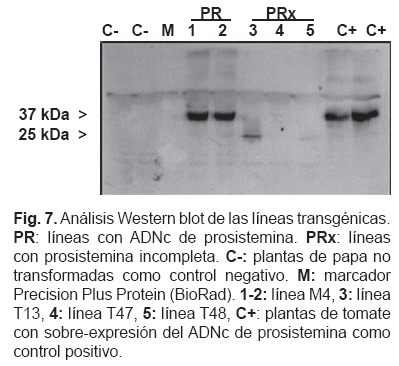
Para controlar si la proteína prosistemina de tomate era producida por las plantas transgénicas, se realizó un análisis Western blot usando el anticuerpo apropiado, la anti-prosistemina (Delano et al., 1999) que se liga a la prosistemina completa. Un ejemplo del Western blot con algunas líneas transgénicas se muestra en la fig. 7.
Una banda de proteína con un peso molecular aparente de 37 kDa se detectó en controles positivos de tomate (McGurl et al., 1994) y en líneas PR portadoras del ADNc de prosistemina (fig. 7), lo cual es diferente al peso molecular real de 23 kDa. Esta migración inesperada ya fue observada previamente en mutantes de prosistemina producidos por E. coliyfueexplicadadebidoalcarácteraltamente hidrofílico de la prosistemina, lo cual modifica su movilidad (Delano et al., 1999). Una banda tenue de aproximadamente 25 kDa también se observó en las líneas PRx. Ninguna de las dos bandas de proteínas se observó en plantas de papa no transformada.
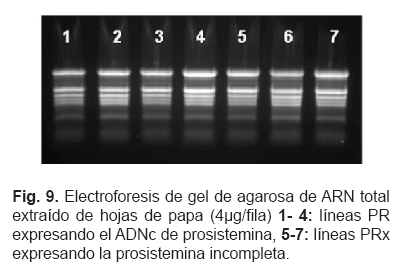
Plantas transgénicas mostraron un efecto específico en la expresión de genes de defensa. Para el análisis de expresión génica las plantas se obtuvieron a partir de tubérculos y las hojas se colectaron a las 4 semanas. Los genotipos PR y PRx se analizaron conjuntamente con plantas de papa no transformadas usadas como control. Se seleccionaron 3 líneas PR y PRx. El fenotipo de las plantas transgénicas (fig. 8) no mostró ninguna diferencia visible comparado con el de las plantas no transformadas.

El ARN total extraído de hojas de 27 plantas, 3 plantas por cada línea transgénica, se analizaron mediante RT-PCR en tiempo real. La electroforesis de gel de agarosa de ARN total de algunas muestras se observa en la fig. 9.
Después de la transcripción inversa delARNm, la calidad del ADNc se controló mediante RT-PCR con los primers Elongation Factor 1-a para revelar cualquier contaminación de ADN. Un ejemplo de electroforesis de gel de agarosa de RT-PCR con los primers EF1-a se observa en la fig. 10.
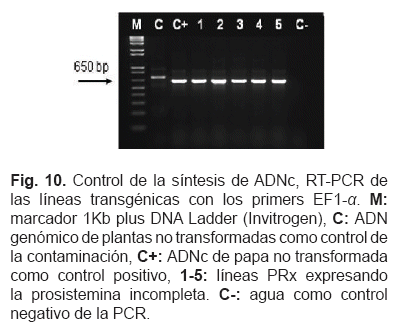
La transcripción de los transgenes se controló de nuevo mediante RT-PCR con los primers Prosys y Sysdel (no mostrado). El gen Elongation Factor 1-alpha se usó como una referencia endógena para normalizar el nivel de expresión de los genes analizados, dado que mostró tener un nivel consistente en todas las muestras (Nicot et al., 2005). Plantas no transformadas se usaron como calibrador. Los valores Ct se usaron para calcular la cantidad relativa (valores RQ) de cada gen en estudio, usando la formula 2"AACt. Los productos de la PCR también se controlaron en gel de agarosa al 2% (no mostrado). Durante el análisis de las curvas de disociación, se confirmó que un único producto fue amplificado y que las temperaturas Tm correspondían al valor calculado en la tabla 2.
Tres grupos de genes se analizaron mediante RT-PCR en tiempo real, todos relacionados con el mecanismo de defensa de la planta. El primer grupo formado por calmodulina (CaM), hidroperoxido l i asa (HPL), y lipoxigenasa 3 (Lox3), todos genes asociados al inicio de la vía de los octadecanoides. CaM y Lox involucrados con la generación de acido 13-hidroperoxi-octadecatrienoico y subsecuente producción de 12-OPDA, componente esencial para la generación de ácido jasmónico, y el gen HPL esencial para la generación de VOCs. El segundo grupo de genes incluyó algunos relacionados al ataque de patógenos, el gen pathogenesis-relatedprotein1b (PR1b) y endo-1,3-beta-D-glucanasa (GluB2). El último grupo consistió en las prosisteminas endógenas de papa 1 y 2 (PotProsys) y los glicopéptidos endógenos o prosisteminas ricos en hidroxipolina (ProHypSys), todos involucrados en la respuesta de la planta a la masticación.
La expresión génica de CaM y HPL no mostraron diferencias en las líneas transgénicas respecto a los controles no transformados (fig. 11). Por el contrario, el gen Lox3 se sobre-expresó 11 veces en las líneas PR y 6 veces en las líneas PRx con respecto a las plantas no transformadas usadas como control (DES). La expresión de los genes entre las líneas transgénicas PR y PRx no mostraron diferencias importantes.

Estos resultados sugieren que la terminal N de prosistemina también juega un rol en la activación de algunos genes de defensa en papa, así como Dombrowski y otros (1999) encontraron que la terminal C es importante en la inducción de inhibidores de proteasas. La sobre expresión de Lox3 en ambas líneas transgénicas sugiere la inducción de algunos genes que participan al inicio de la cascada de eventos.
De los genes relacionados con la respuesta a patógenos, el gen PR1b mostró alguna inhibición en las líneas PRx, y GluB2 tuvo una expresión diferente en las líneas PR comparadas con el control, aun así, después del análisis estadístico los datos no mostraron diferencias significativas (fig. 12).
Las dos prosisteminas endógenas de papa (PotProsys 1 y 2) se analizaron juntas debido a la semejanza de sus secuencias. La prosistemina de papa y los glicopéptidos ricos en hidroxiprolina (péptidos similares a la prosistemina), ambos relacionados con la defensa, no mostraron diferencias en el nivel de expresión en ambas líneas transgénicas en comparación con el control no transformado, sugiriendo que la prosistemina de tomate no produce una activación de estos genes específicos de papa (fig. 13).
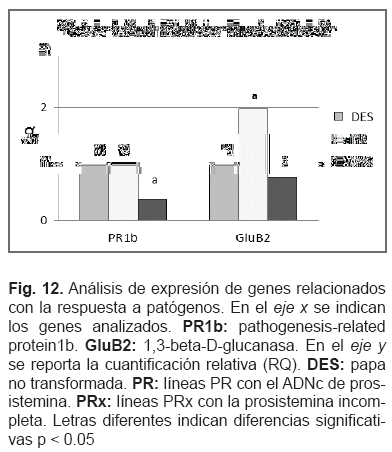
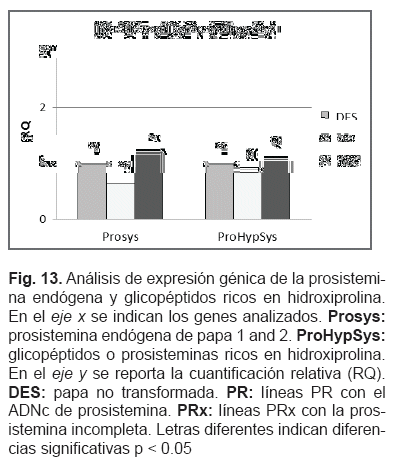
Previamente, se demostró que la sistemina del tomate, un potente activador de la vía de los octadecanoides, produce una activación sistémica de genes de defensa mediante la amplificación del acido jasmónico en tomate (Li et al, 2002; Ryan y Pearce, 2003; Corrado et al, 2007). Cuando se expresa en papa, la sistemina parece provocar un efecto específico. Todos los genes analizados tienen relación con la defensa de la planta, pero solo Lox3 fue sobre-expresado. Halitschke y Baldwin, (2003) demostraron en Nicotiana attenuata que Lox3 provee de acidos grasos hidroperóxidos para la síntesis de jasmonatos, el cual es una señal clave para la transcripción de proteínas defensivas. Este planteamiento coincide con Narváez-Vásquez y Ryan (2002) quienes observaron que la expresión del ADNc de prosistemina en papa regula la síntesis de inhibidores de proteasas en hojas. Así, estos datos sugieren que la prosistemina de tomate provoca en papa, de modo parcial, un efecto similar al que provoca en tomate. Las dos líneas transgénicas de papa mostraron una sobre expresión en Lox3, indicando que la terminal N de la proteína prosistemina juega un rol en la activación de Lox3.
CONCLUSIONES
La expresión del gen prosistemina en papa, no afecta los genes relacionados con respuesta a patógenos como GluB2 y PR1b. Los genes de las prosisteminas endógenas relacionadas con el mecanismo de defensa, Prosys y ProHypSys no fueron afectados por el transgen. Solamente Lox3 se sobre expresó en ambos transformantes 11 veces más, lo cual indica que la terminal N también contribuye en la activación de algunos genes de defensa en papa. El estudio demuestra que la expresión heteróloga de precursores de una molécula señal puede provocar efectos inesperados, en este caso, potencialmente útiles para fines biotecnológicos.
AGRADECIMIENTOS
Los autores agradecen al Dr. Giandomenico Corrado por sus valiosos consejos y constante apoyo, al Prof. Teodoro Cardi por sus sugerencias en la transformación genética de papa y al Prof. Andreas Schaller por suministrar el antisuero de prosistemina.
REFERENCIAS
Beerhues L. and Kombrink E., 1994. Primary structure and expression of mRNAs encoding basic chitinase and 1,3-beta-glucanase in potato. Plant Molecular Biology 24(2): 353-367. [ Links ]
Bhattacharya R., Pearce G., Ryan C.A., 2008. A polyprotein precursor of defense-related hydroxyproline-rich glycopeptides in Solanum tuberosum. Unpublished. [ Links ]
Bradford M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254. [ Links ]
ChenH.,WilkersonC.G.,KucharJ.A.,Phinney B.S., HoweA.W., 2005. Jasmonate-inducible plant enzymes degrade essential amino acids in the herbivore midgut. PNAS, USA 102(52): 19237-19242. [ Links ]
Constabel C.P., Lynn Y. and Ryan C.A., 1998. Prosystemin from potato, black nightshade, and bell pepper: primary structure and biological activity of predicted systemin polypeptides. Plant Molecular Biology 36: 52-62. [ Links ]
Corrado G., Sasso R., Pasquariello M., Iodice L., Carretta A., Cascone P., Ariati L., Digilio M.C., Guerrieri E., Rao R., 2007. Systemin regulates both systemic and volatile signaling in tomato plants. Journal of Chemical Ecology 4: 669-681. [ Links ]
Delano J.P., Dombrowski J.E., Ryan C.A., 1999. The expression of tomato prosystemin in Escherichia coli: a structural challenge. Protein Expression and Puri fication 17: 74-82. [ Links ]
DombrowskiJ.E.,PearceG.,RyanC.A.,1999. Proteinase inhibitor-inducing activity of the prohormone prosystemin resides exclusively in the C-terminal systemin domain. PNAS, USA 96 (22): 12947-12952. [ Links ]
Felton G.W., 2005. Indigestion is a plant's best defense. PNAS, USA 102(52): 18771-18772. [ Links ]
Halitschke R. and Baldwin I.T., 2003. Antisense LOX expression increases herbivore performance by decreasing defense responses and inhibiting growth-related transcriptional reorganization in Nicotiana attenuata. The Plant Journal 36: 794-807. [ Links ]
Hanley M.E., Lamont B.B., Fairbanks M.M., Rafferty C.N., 2007. Plant structural traits and their role in anti-herbivore defence. Perspectives in Plant Ecology, Evolution and Systematics 8: 157-178. [ Links ]
Hoegen E., Stroemberg A., Pihlgren U. and Kombrink E., 2002. Primary structure and tissue-specificexpression of the pathogenesis-related protein PR-1b in potato. Mol. Plant Pathol. 3(5): 329-345. [ Links ]
Howe G.A., Jander G., 2007. Plant immunity to insect herbivores. Annual Review of Plant Biology 59: 41-66. [ Links ]
Kolomiets, M.V., Hannapel, D.J. and Gladon, R.J., 1996. Nucleotide Sequence of a cDNA Clone for a Lipoxygenase from Abscisic Acid-Treated Potato Leaves (Accession No. U60202). Plant Physiology 112: 445. [ Links ]
Livak K.L., Schmittgen T.D., 2001. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2"AACt method. Methods 25: 402-408. [ Links ]
Li L., Li C., Lee G.I., Howe G.A., 2002. Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato. PNAS, USA 99: 6416-6421. [ Links ]
McGurl B., Ryan C.A., 1992. The organization of the prosystemin gene. Plant Molecular Biology 20: 405-409. [ Links ]
McGurl B., Pearce G., Orozco-Cárdenas M., Ryan C.A., 1992. Structure, expression, and antisense inhibition of the systemin precursor gene. Science 255: 1570-1573. [ Links ]
McGurl B., Orozco-Cárdenas M., Pearce G., Ryan C.A., 1994. Overexpression of the prosystemin gene in transgenic tomato plants generates a systemic signal that constitutively induces proteinase inhibitor synthesis. PNAS, USA 91: 9799-9802. [ Links ]
Nakane E., Kawakita K., Doke N., Yoshioka H., 2003. Elicitation of primary and secondary metabolism during defense in the potato. Journal of General Plant Pathology 69: 378-384. [ Links ]
Narváez-Vásquez J. and Ryan C.A., 2002. The systemin precursor gene regulates both defensive and developmental genes in Solanum tuberosum. PNAS, USA 99: 15818-15821. [ Links ]
Narváez-Vásquez J., Orozco-Cárdenas M.L., Ryan C.A., 2007. Systemic wound signaling in tomato leaves is cooperatively regulated by systemin and hydroproline-rich glycopeptide signals. Plant Mol Biol. 65: 711-718. [ Links ]
Nicot N., Hausman J.F., Hoffmann L., Evers D., 2005. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. Journal of Experimental Botany 56(421): 2907-2914. [ Links ]
Pearce G., Strydom D., Johnson S., Ryan C.A., 1991. A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science 253(5022): 895-897. [ Links ]
Pearce G., Ryan C.A., 2003. Systemic signalling in tomato plants for defense against herbivores. Isolation and characterization of three novel defense-signaling glycopeptides hormones coded in a single precursor gene. The Journal of Biological Chemistry 278(32): 30044-30050. [ Links ]
PearceG., SiemsW.F., BhattacharyaR., Chen Y.C., Ryan C.A., 2007. Three hydroxyproline-rich glycopeptides derived from a single petunia polyprotein precursor activate defensin I, a pathogen denfense response gene. The Journal of Biological Chemistry 284(24): 17777-17784. [ Links ]
PearceG., BhattacharyaR., Chen Y. C.,Barona G., Yamaguchi Y., Ryan C.A., 2009. Isolation and characterization of hydroxyproline-rich glycopeptides signals in black nightshade leaves. Plant Physiology 150(3): 1422-1433. [ Links ]
Rotenberg D., Thompson T.S., German T.L., Willis D.K., 2006. Methods for effective real-time RT-PCR analysis of virus-induced gene silencing. Journal of Virological Methods 138: 49-59. [ Links ]
Ryan C.A., 2000. The systemin signaling pathway: differential activation of plant defensive genes. Biochimica et Biophysica Acta 1477: 112-121. [ Links ]
Ryan C. A. y Pearce G., 2003. Systemins: a functionally defined family of peptide signals that regulate defensive genes in Solanaceae species. PNAS, USA 100: 14577-14580. [ Links ]
Sambrook J. and Russell D.W., 2001. Molecular Cloning: a laboratory manual 3rd edition. Cold Spring Harbor Laboratory Press pp. 999. [ Links ]
Takezawa D., Liu Z.H., An G., Poovaiah B.W., 1995. Calmodulin gene family in potato: developmental and touch-induced expression of the mRNAencoding a novel isoform. Plant Mol. Biol. 27(4): 693-703. [ Links ]
VancanneytG., SanzC., FarmakiT., Paneque M., Ortego F., 2001. Hydroperoxyde lyase depletion in transgenic potato plants leads to an increase in aphid performance. PNAS, USA 98: 8139-8144. [ Links ]
Walling L.L., 2000. The myriad plant responses to herbivores. Journal of Plant Growth Regulation 19: 195-216. [ Links ]