








Services on Demand
Article
 Article in pdf format
Article in pdf format Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
 Cited by SciELO
Cited by SciELO
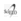 Similars in SciELO
Similars in SciELO
Bookmark
Revista Científica Agrociencias Amazonía
Print version ISSN 2307-9606
Rev.Cient.Agro.Amaz. vol.1 no.1 Beni May 2013
ARTICULO
Cambio Estacional de la Estructura de Comunidades Ictícolas en Relación a la Conectividad y Cobertura Vegetal en Pozas Artificiales en la Amazonia Boliviana
Yunoki T.*, Cholima B. R., Moreno J. P. R., Jiménez J. V. y Torres L. V.
Centro de Investigación de Recursos Acuáticos (CIRA), Universidad Autónoma del Beni "José Ballivián", Trinidad, Beni, Bolivia. *E-mail de contacto: takayukiyunoki@yahoo.com
RESUMEN
El presente estudio describe el cambio estacional de la estructura de comunidades ictícolas en relación a la conectividad y la cobertura vegetal (CV) en pozas artificiales de la llanura de Mojos, así como sugiere los procesos ecológicos que influyen en las diferentes escalas espacio - temporales. Se recolectaron muestras mensuales durante dos ciclos hidrológicos del 2009 a 2011. El muestreo se realizó en dos pozas aisladas y dos pozas conectadas durante el primer año, e igualmente durante el segundo año de estudio pero en diferentes pozas, abarcando así 4 tratamientos diferentes al año. El análisis de correspondencia segmentado (DCA) muestra la organización jerárquica, donde la conectividad mediante la asociación de las estrategias de historia vital ejerce control a escala espacial mayor entre las pozas conectadas y aisladas, mientras la CV mediante la adaptación a la respiración aérea ejerce control a escala espacial menor dentro de las pozas conectadas o aisladas con la bajada de aguas. Durante la inundación anual se observa la homogenización de las estructuras de comunidades ictícolas debido a la disminución de peces y la dominancia de sardinas muy pequeñas. La asociación de las tres estrategias de historia vital a la heterogeneidad espacial originada por la conectividad no es permanente durante el ciclo hidrológico. Los peces pequeños, probablemente asociados a la estrategia oportunista, colonizan todas las pozas durante la inundación anual pero no persisten durante la época seca donde la tasa de depredación es alta. Los peces grandes, probablemente asociados a la estrategia epocanista, requieren de la inundación anual y persisten durante la época seca.
Palabras claves: estrategia de historia vital, inundación anual, estructura de comunidades icticolas
ABSTRACT
The present study describes the seasonal change of fish assemblage structure related to the connectivity and the vegetation cover (VC) in artificial ponds of Mojos plain, also suggests the ecological processes that operate in different spatial - temporal scales. Monthly samples were collected during two hydrologic cycles between 2009 and 2011. Samples were collected in two isolated ponds and two connected ponds during the first year; the second year, samples were collected from another two isolated and two connected ponds. Four different treatments each year. The Detrended Correspondence Analysis (DCA) shows the hierarchical organization where the connectivity by means of the association of life history strategies controls in large spatial scale between the connected ponds and also isolated ponds. Moreover, the VC through the adaptation to the aerial breathing controls in small spatial scale among the connected or isolated pools during water decrease. Evenmore, during annual flood, the homogenization of fish assemblage structures was observed, due to the reduction of fish and dom inance of very small tetras. The association of the three life history strategies to the spatial heterogeneity originated by connectivity is not permanent during the water cycle. Small fish probably associated to the opportunistic strategy, colonize all ponds during annual flood, but do not persist during dry season where predation rate is high. Large fish, probably associated to the periodic strategy, require the annual flood and they persist during dry season.
Keywords: life history strategy, annual flood, fish assemblage structure
INTRODUCCIÓN
El sector pesquero está obligado a explotar nuevas especies alternativas, así mismo adquirir un moderno sistema de control introduciendo nuevas técnicas de proliferación en el medio natural y el cultivo en cautiverio. Frecuentemente la comunidad científica, así como las autoridades regionales orientan su interés al control y cultivo en cautiverio para especies comerciales, pero raramente a la explotación de nuevas especies alternativas, menos aún a su proliferación en el medio natural. Considerando el potencial pesquero de las especies comerciales en el medio natural, así como el costo de producción en cautiverio, la política pesquera actual no está siendo orientada directamente a la seguridad alimentaria y el alivio de la pobreza.
La llanura de Mojos es pobre en nutrientes, heterogénea, y con una dinámica estacional (Euroconsult, 1999). El pulso de inundación es el mayor promotor de este ecosistema, ya que las aguas transportan nutrientes como materias disueltas suspendidas orgánicas e inorgánicas y estos nutrientes son indispensables para los seres vivos del ecosistema (Junk et al., 1989). La mayoría de los peces aprovecha los diferentes medios y migra estacionalmente (epocanistas), mientras la minoría aprovecha de los medios particulares o efímeros, siendo así sedentarios (equilibristas) o colonizadores (oportunistas) (Winemiller, 1989).
Al retroceder las aguas los organismos acuáticos, sobre todos los peces, se concentranencuerposdeaguas permanentes.
En el área de influencia de Trinidad se observan numerosas pozas artificiales, en algunas de las cuales la población de los peces es mucho mayor que en cuerpos de aguas permanentes y naturales, por ejemplo los bucheres (Hoplosternum littorale). Esto nos recuerda la cultura de Mojos y el control que tenían sobre el agua, una extensa obra civil que proveía de refugio a los habitantes durante las inundaciones y el suministro de alimentos permitiendo, mediante el cultivo de productos agrícolas (Lee, 1996). El control hidráulico dirigido al suministro de alimentos a través de una eficiente recolección de la producción alóctona de la llanura de Mojos, era un sistema de producción abierto al sistema fluvial que requería de algunos trabajos para contrarrestar la eutrofización y sedimentación, y así recolectar peces y taropé.
Este estudio comparativo sustenta el sistema de producción de la cultura de Mojos, probando los efectos de dos factores cada uno con dos categorías: 1. Conectividad con la zona de inundación (aislada o conectada), 2. Cobertura vegetal del taropé (cubierta o descubierta) sobre la estructura de las comunidades ictícolas en pozas artificiales en el área de influencia de Trinidad, durante la época seca. Se conoce que, la biomasa es mayor en las pozas conectadas, sobre todo en las pozas cubiertas sobrepasando 1 kg/ m2 de espejo de agua hasta 3 kg/m2. Las especies de estas pozas poseen mayor porte corporal y están adaptadas a la respiración aérea, así estas especies están asociadas a la estrategia vital epocanista para aprovechar una expansión anual de producción acuática. Durante la época seca se refugian en donde la baja concentración del oxígeno disuelto y la cobertura vegetal ofrece protección de peces depredadores como palometa (Pygocentrus nattereri), surubí (Pseudoplatystoma spp.), etc., y de aves depredadoras como son cuervo, garza, bato, etc. (Yunoki et al., 2012).
Los grandes ríos de la región neotropical presentan un ciclo anual de inundación predecible, por ejemplo en las lagunas del río venezolano Orinoco, la inundación anual homogeniza las estructuras de comunidades ictícolas, la diferencia ocurre al inicio de la época seca, disminuyendo las especies visualmente orientadas respecto a las especies adaptadas a baja visibilidad (Rodríguez y Lewis, 1994). La comunidad de peces está organizada por piscívoros bajo la influencia de la transparencia controlada por la profundidad y área, este proceso ecológico se denomina modelo Piscívoro de Transparencia Morfológica (PTM) (Rodríguez y Lewis, 1997). En las lagunas del río Mamoré Central de la Amazonía boliviana, la conectividad limita los movimientos de los peces entre las lagunas de planicie y los cauces abandonados. El modelo PTM ejerce mayor control en los cauces abandonados, donde la limitación de movimientos es débil. Así, la diferencia entre las lagunas de planicie y los cauces abandonados puede interpretarse de las asociaciones de estrategias de historia vital. El acceso estacional del río Mamoré y la heterogeneidad espacial originada por la conectividad influirán en la colonización y la persistencia (Pouilly y Rodríguez, 2004).
La ecología moderna pretende explicar la distribución de las comunidades, integrando los procesos ecológicos que intervienen en las diferentes escalas espacio-temporales. El estudio comparativo es de naturaleza instantánea y los estudios estacionales, como los casos de las lagunas, sugieren inferencias sobre los procesos ecológicos. El presente estudio describe el cambio estacional de las estructuras de comunidades ictícolas en relación a la conectividad y la cobertura vegetal, así como, sugiere los procesos ecológicos que intervienen en las diferentes escalas espacio-temporales en las pozas artificiales de la llanura de Mojos. Mediante el presente estudio se desea verificar si la conectividad limita el movimiento de peces entre las pozas aisladas y conectadas durante la inundación anual; además, comprobar si las especies de pozas aisladas están asociadas a la estrategia de historia vital "oportunista" o "equilibrista", las cuales no aprovechan la heterogeneidad de medios ni la dinámica estacional, por otro lado, las especies de pozas conectadas estarán asociadas a la estrategia de historia vital "epocanista", con aprovechamiento de la expansión anual de producción acuática, conociendo además que, la asociación entre las estrategias de historia vital y la heterogeneidad espacial originada por la conectividad es permanente durante el ciclo hidrológico.
MATERIALES Y MÉTODOS
Área de estudio
La llanura amazónica del Mamoré Central tiene un clima tropical, con la estación de lluvias concentrada en verano. Del 60 a 80% de las precipitaciones ocurre entre diciembre y marzo al mismo tiempo que las temperaturas son las más altas del año. Los niveles de aguas más importantes son registrados generalmente entre los meses de enero y marzo (Roche et al., 1992). Los llanos de Bolivia forman en tiempo de aguas altas una inmensa zona inundada de bosques y pampas, que se encuentra entre las más extendidasyproductivasdelmundo(Lauzanne et al., 1990). Los diversos medios acuáticos del Mamoré Central están bajo influencia de las aguas cuyas características están ligadas a su origen, así las del río Mamoré originadas en losAndes son relativamente mineralizadas, por otra parte, las aguas de planicie más directamente ligadas a la pluviosidad local tienen una mineralización baja (Corbin et al., 1988). El área de estudio corresponde a la zona inundada de la pampa, donde las aguas están más sujetas a la pluviosidad local. La zona presenta principalmente lagunas poco profundas de forma rectangular, los ríos menores, el rastro de camellones construidos por la civilización ancestral (Lee, 1996) y las numerosas pozas artificiales construidas por la civilización moderna por diversas razones como aplanar el terreno, construcción de carreteras, aguadas para consumo humano o animal, etc.
Muestreo
Durante el estudio comparativo se recolectaron las muestras en 12 pozas artificiales durante la época seca, en septiembre 2010, para determ inar el efecto de la conectividad en la zona de inundación (conectada o aislada) y la cobertura vegetal de taropé (cubierta o descubierta) sobre las estructuras de comunidades ictícolas. El diseño experimental utilizado fue 2 por 2 factorial (Yunoki et al., 2012).
Para el estudio estacional se recolectaron muestras mensuales en el periodo de dos ciclos hidrológicos. El muestreo se realizó en 2 pozas aisladas y 2 pozas conectadas durante el primer año (julio 2009 a junio 2010) y otras 2 pozas aisladas y 2 pozas conectadas durante el segundo año (octubre a noviembre 2010, enero a septiembre 2011 y diciembre 2011), con 4 tratamientos diferentes por año, correspondientes a las pozas AC1, AD1, CC1, CD1 yAC2, AD2, CC2, CD2 (figura 1).
La recolección de peces cubrió aproximadamente el 20% de la superficie de la poza con una malla de arrastre de 5 mm de rombo entre nudo a nudo, 60 m de largo y 4 m de alto. Para calcular el área cubierta se registró el largo (m) y ancho (m) en ambos extremos. Todos los especímenes fueron conservados en hielo o formol al 10% y luego trasladados al laboratorio. Los especímenes de referencia (tabla 1) fueron conservados en el Museo Ictícola de la Universidad Autónoma del Beni.
Análisis de datos
Las comunidades ictícolas fueron expresadas en abundancia de especie/1 00m2 de espejo de agua, y luego transformadas por logaritmo natural (abundancia+1). Las especies con abundancia menor al 5% fueron excluidas del análisis posterior. Se aplicó el análisis de correspondencia segmentado (DCA) para ilustrar el cambio estacional de las estructuras de comunidades ictícolas mediante los trayectos de sitios en su ordenación. El DCA fue realizado por el programa R con sus paquetes Permute 0.7-0y Vegan 2.0-5.

RESULTADOS
Abundancia y Biomasa
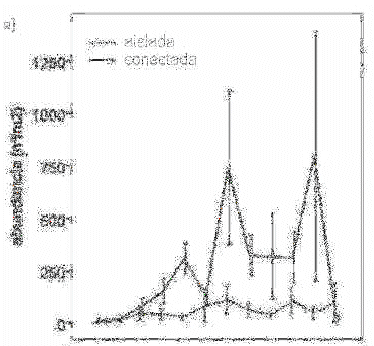
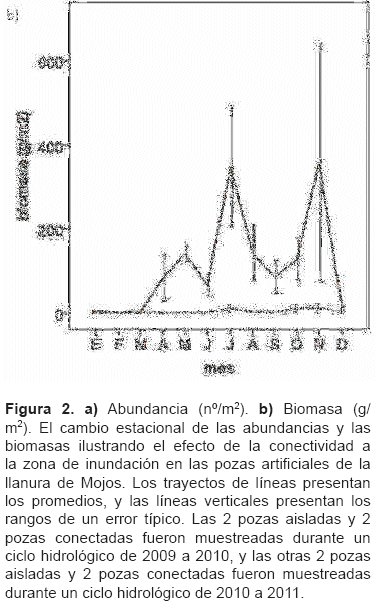
La abundancia (n°/m2) y, sobre todo, la biomasa (g/m2) de las pozas conectadas, muestran mayor fluctuación estacional a lo largo del ciclo hidrológico. Se observa la disminución brusca con el inicio de la subida de aguas en el mes de diciembre, y se mantiene en baja durante la inundación anual de enero a marzo. Se observa el incremento gradual con la bajada de aguas de abril a julio (figura 2a y 2b).
Comunidades ictícolas
Se recolectaron 72 especies de peces que correspondieron a 54517 individuos y 1720,7 kg,delascuales38especies(52,8%)superaron a 5% en el número de individuos de algunas muestras, estas especies correspondieron a 53610 individuos (98,3%), y 1703,6 kg (99%) de la captura total, mostrados en la tabla 1.
La ordenación en los dos primeros ejes de DCA muestra que los trayectos estacionales de los sitios del primero y segundo año fueron similares, si la conectividad y la CV pertenecen a la misma categoría. Los trayectos estacionales se sobreponen durante la inundación anual, más que todo, de febrero a marzo, y se apartan gradualmente con la bajada de aguas desde abril hasta el inicio de la subida de aguas en diciembre - enero, reflejando sus pertenencias categóricas de la conectividad y la CV. Los trayectos de pozas conectadas son similares en diagonal de derecha superior a izquierda inferior. La diferencia entre las pozas conectadas despejadas y cubiertas, se evidencia con el progreso de la época seca en diagonal de derecha inferior a izquierda superior. Los trayectos de pozas aisladas despejadas no son prominentes. Por último los trayectos de pozas aisladas cubiertas van de derecha inferior a izquierda superior (figura 3a).


La sobreposición de los trayectos durante la inundación anual, o la homogenización de las estructuras de comunidades ictícolas, se debe a la disminución de peces con una dominancia de Sardinas muy pequeñas como Ctenobrycon spilurus y Serrapinnus piaba. Los trayectos posterior de pozas conectadas se deben al incremento de los peces de mayor porte corporal con la adaptación a la respiración aérea como Buchere (Hoplosternum littorale) y Yayú (Hoplerythrinus unitaeniatus), y la diferencia entre las pozas conectadas despejadas y cubiertas con el progreso de la época seca se debe a la dominancia de las especies no adaptadas a la respiración aérea como Torito (Trachelyopterus galeatus), Cuchillo (Eigenmannia virescens), y Sardina pequeña (Gymnocorymbus ternetzi) relativas a las especies adaptadas a la respiración aérea como Buchere (Megalechis picta, Callichthys callichthys), Cuchillo (Brachyhypopomus brevirostris) y Yayú (Erythrinus erythrinus). Aúnque no se aprecia en la ordenación, los peces piscívoros como la Palometa (Pygocentrus nattereri) están presentes en las pozas conectadas despejadas. Los trayectos de pozas aisladas despejadas no son prominentes, lo cual se debe a la dominancia permanente de Sardinas muy pequeñas ya anteriormente citadas como también la Serepapa amarilla (Bujurquina cf. vittata). Los trayectos de pozas aisladas cubiertas se deben a la dominancia de especies adaptadas a la respiración aérea como Anguilla (Synbranchus madeirae) y Pez pulmonado (Lepidosiren paradoxa) con cuerpos muy largos y Pez anual (Rivulus beniensis) (figura 2a y 2b, figura 3b).
DISCUSIÓN
La ecología moderna pretende explicar la distribución de comunidades, integrando los procesos ecológicos que ejercen en las diferentes escalas espacio - temporales (Rodríguez y Lewis, 1994, 1997, Pouilly y Rodríguez, 2004). En el presente estudio, la conectividad presume establecer la heterogeneidad espacial en cuanto a la variabilidad de factores ambientales, abundancias de recurso y tasas de depredación, lo cual ejerce el control mediante la asociación de las estrategias de historia vital en la escala espacial mayor, entre las pozas aisladas y conectadas, y la CV ejerce el control en la escala espacial menor mediante la adaptación a la respiración aérea, dentro de las pozas aisladas o conectadas con la bajada de aguas.
Durante la inundación anual ocurre la homo-genización de las estructuras de comunidades ictícolas debido a la disminución de peces con una dominancia de Sardinas muy pequeñas, y las pozas aisladas se establecen a una baja densidad poblacional mientras las pozas conectadas se establecen a una alta densidad poblacional al inicio de la época seca. Con el progreso de la época seca, las especies adaptadas probablemente a la estrategia oportunista incrementan la densidad poblacional en las pozas aisladas puesto que estas pronto alcanzaran el tamaño de madurez sexual en un ambiente vacio, mientras se extinguen en las pozas conectadas puesto que la tasa de depredación es fuerte para estas especies. Las especies adaptadas probablemente a la estrategia epocanista no incrementan la densidad poblacional en las pozas aisladas ya que habrían pasado la época de reproducción, tampoco no alcanzaran el tamaño de madurez sexual, mientras incrementan la densidad poblacional en las pozas conectadas debido a la disminución brusca de espejos de aguas y menor tasa de depredación para estas especies de porte mayor. Lo que se observa con el progreso de la época seca es la dominancia de especies muy pequeñas probablemente adaptadas a la estrategia oportunista en las pozas aisladas, y la dominancia de especies de porte mayor, probablemente adaptadas a la estrategia epocanista en las pozas conectadas. Considerando la baja biomasa, se presume la baja tasa de depredación en las pozas aisladas, así la cobertura vegetal y la adaptación a respiración aérea no implicarían la interacción presa predador, sino la adaptación a respiración aérea y los largos cuerpos de Anguilla y Pez pulmonado parecerían permitir el movimiento sobre tierras, explotando los recursos distribuidos entre diferentes pozas. Considerando la gran biomasa, se presume la alta tasa de depredación en las pozas conectadas. Aquí la cobertura vegetal y la adaptación a respiración aérea implicarían la interacción presa predador, sea indirecta como la fuga de depredadores o directa como la mortalidad causada por depredación. En las pozas conectadas dominan las especies adaptadas a la estrategia epocanista como se espera en otros cuerpos de aguas permanentes en el medio natural, la mayor concentración de peces en las pozas cubiertas de taropé podría deberse a la fuga de los peces depredadores como Palometa (Pygocentrus nattereri), Surubí (Pseudoplatystoma spp.), etc., y las aves depredadoras como Cuervo, Garza, Bato, etc.
El presente estudio observó lo contrario a las hipótesis planteadas. La inundación anual promueve la homogenización de las estructuras de comunidades ictícolas, además la asociación de las tres estrategias de historia vital a la heterogeneidad espacial, originada por la conectividad, no es permanente durante el ciclo hidrológico.
La teoría de estrategias de historia vital supone que las poblaciones o las especies se ajustan a la relación entre tácticas demográficas-reproductivas para maximizar la adaptabilidad al medio donde habitan, como resultado de la selección natural. Existen teorías de estrategias de historia vital, entre las más conocidas son una compuesta de dos estrategias (Pianka, 1970) y otra compuesta de tres estrategias (Winemiller, 1989). En este último, la teoría se originó del estudio empírico, en lo cual analiza la relación entre las 10 tácticas demográficas reproductivas de 71 especies, y la asociación de las tres estrategias de historia vital a los tres hábitat diferentes en el paisaje; río en el pie de monte Andino, río en las tierras bajas, y represa, en el llano Venezolano (Winemiller, 1989). Las tres estrategias pueden ser clasificadas como, 1. Los peces pequeños, madurez temprana y cortas vidas (oportunista), 2. Los peces grandes, altas fecundidades y largas vidas (epocanista), y 3. Los peces medianos que exhiben los cuidados paternales, y producen menor número y mayor tamaño de huevos (equilibrista) (Winemiller & Rose, 1992).
Las tres estrategias se originaron del estudio empírico de las comunidades ictícolas de hábitat diferentes, sin embargo, estas estrategias no son mutuamente exclusivas (Winemiller, 1989). Las tres estrategias mediante algún proceso ecológico, en menor escala espacio - temporal, persistirían. El presente estudio percibe las tres estrategias de historia vital en un conjunto de las comunidades ictícolas interconectadas por dispersión y sugiere las inferencias sobre el proceso ecológico. Los peces pequeños probablemente asociados a la oportunista colonizan todas las pozas durante la inundación anual sin embargo no persisten durante la época seca donde la tasa de depredación es alta. Los peces grandes probablemente asociados a la epocanista requieren de la inundación anual y persisten durante la época seca donde la tasa de depredación es alta.
CONCLUSIÓN
El cambio estacional de las estructuras de comunidades ictícolas en relación a la conectividad y la cobertura vegetal (CV) en las pozas artificiales de la llanura de Mojos muestra una organización jerárquica, donde la conectividad mediante la asociación de las estrategias de historia vital ejerce control en la escala espacial mayor entre las pozas conectadas y aisladas, mientras la CV mediante la adaptación a la respiración aérea ejerce control en la escala espacial menor adentro de las pozas conectadas o aisladas con la bajada de aguas. Durante la inundación anual se observa la homogenización de la estructura de comunidades ictícolas debido a la disminución de peces y la dominancia de Sardinas muy pequeñas. La asociación de las tres estrategias de historia vital a la heterogeneidad espacial originada por la conectividad no es permanente durante el ciclo hidrológico. Los peces pequeños probablemente asociados a la oportunista colonizan todas las pozas durante la inundación anual sin embargo no persisten durante la época seca donde la tasa de depredación es alta. Por el contrario, los peces grandes probablemente asociados a la epocanista requieren de la inundación anual y persisten durante la época seca.
AGRADECIMIENTOS
Los autores expresamos nuestro agradecimiento a Marc Pouilly, Institut de Recherche pour le Développement (IRD), y Pascal I. Hablützel, KU Leuven, Laboratory for Animal Diversity and Systematics por el constante apoyo técnico científico. De igual manera a Alfredo Parada Montalbán, a todo el personal del Modulo de Piscicultura de la Facultad de Ciencias Agrícolas y a los estudiantes de las Facultades de Ciencias Pecuarias y Agrícolas por el constante apoyo.
REFERENCIAS BIBLIOGRÁFICAS
Corbin, D., J. Guyot, H. Calle y J. Quintanilla (1988) Datos físico químicos de los medios acuáticos de la zona del Mamoré Central, Región de Trinidad, Amazonía Boliviana, Informe Científico N° 8, Convenio ORSTOM, U.T.B. CORDEBENI, Trinidad, Bolivia, 58 p. [ Links ]
Euroconsult, CGL Consultores Galindo (1999) Zonificaciónagroecológicaypropuestatécnica del plan de uso del suelo del departamento de Beni. Programa para el Ordenamiento Territorial de la región amazónica boliviana en los departamentos de La Paz, Beni y Cochabamba: 216 p [ Links ]
Junk W. J., P.B. Bayley, & R.E. Sparks (1989) The Flood pulse concept in river-floodplain systems, pp. 110-127 in Dodge DP, editor. Proceedings of the International Large River Symposium. Can.Spec.Publ.Fish.Aquat.Sci. [ Links ]
Lee K. (1996) Apuntes sobre las obras hidráulicas prehispánicas de las llanuras de Moxos. Paitití, Año 11, N°1: 24-26, Casa de la Cultura de Beni, Trinidad. [ Links ]
Lauzanne L., G. Loubens y B. Le Gennec (1990) Pesca y biología pesquera en el Mamoré Medio (Región de Trinidad, Bolivia), Interciencia, 15 (6): pp. 452-460 [ Links ]
Pianka ER (1970) On r- and K-selection. Am Nat100: pp. 592-597 [ Links ]
Pouilly M. & M. A. Rodríguez (2004) Determinism of fish assemblage structure in Neotropical floodplain lakes: influence of internal and landscape lake conditions, Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries (LARS2) II: pp. 243-265 [ Links ]
Roche M., C. Fernandez, A. Aliaga, J. Peña, E. Salas, y J. Montaño (1992) Balance hídrico superficial de Bolivia [ Links ]
Rodríguez M. A. & W. M. Lewis Jr. (1994) Regulation and stability in fieh assemblages of neotropical floodplain lakes, Oecologia, 99: pp. 166-180 [ Links ]
Rodríguez M. A. & W. M. Lewis Jr. (1997) Structure of fish assemblades along environmental gradients in floodplain lakes of the Orinoco river, Ecological Monographs, 67(1): pp. 109-128 [ Links ]
Winemiller K.O. (1989) Patterns of variation in life history among South America fishes in seasonal environments, Oecologia, 81: pp. 228-241 [ Links ]
Winemiller K.O. & K. A. Rose (1992) Patterns of life-history diversification in North American fishes: implications for population regulation, Canadian Journal of Fishes and Aquatic Sciences, 14 (10): pp. 2196-2218 [ Links ]
Yunoki T., Cholima B. R. y Moreno J. P. R. (2012) Prospectiva de recolectar peces con el control hidráulico en las llanuras de Mojos (Amazonía Boliviana): Efectos de conectividad y cobertura vegetal sobre la biomasa y la estructura de comunidades ictícolas en las pozas artificiales, Medio Acuático, 3: pp. 1-9 [ Links ]