








Servicios Personalizados
Articulo
 Articulo en PDF
Articulo en PDF Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
Links relacionados
 Citado por SciELO
Citado por SciELO
 Similares en SciELO
Similares en SciELO
Bookmark
BIOFARBO
versión impresa ISSN 1813-5363
BIOFARBO v.17 n.2 La Paz 2009
TRABAJOS DE INVESTIGACIÓN
Optimización de las condiciones de cultivo anaeróbico termófilo para aumentar la producción enzimática de xilanasas con cepas bacterianas encapsuladas y libres
Optimization of anaerobic thermophilic culture conditions for increasing xylanase enzyme production with encapsulated and free bacteria strains
Esther Casablanca Alarcón, Neida,Ríos Manríquez, María Teresa Álvarez, EnriquenTerrazas Siles
Área de Biotecnología, Instituto de Investigaciones Fármaco Bioquímicas, Facultad de Ciencias Farmacéuticas y Bioquímicas, Universidad Mayor de San Andrés. La Paz, Bolivia.
Dirección para correspondencia: Esther Casablanca Alarcón. Instituto de Investigaciones Fármaco Bioquímicas, Facultad de Ciencias Farmacéuticas y Bioquímicas, Universidad Mayor de San Andrés. Av. Saavedra 2224. La Paz, Bolivia.
E mail: sthr_13@hotmail.com
Recibido para publicación en 9/09/09
Aceptado en 20/12/09
RESUMEN
Las bacterias termófilas productoras de enzimas son de gran interés en biotecnología debido a que tienen diversas aplicaciones en la industria por su estabilidad extrema. Estos biocatalizadores pueden ser empleados en la preparación de la pulpa del papel, procesamiento de alimentos agroindustriales y conversión de la hemicelulosa a biomasa. Es por eso que el propósito de este trabajo es optimizar la producción enzimática a fin de obtener mayor actividad xilanolítica de tres cepas bacterianas anaerobias termófilas FT3, 2B y P5 las cuales fueron aisladas del altiplano boliviano. Fue seleccionada la cepa FT3 porque mostró la mayor actividad xilanolítica de 2.0 UI/mL en comparación de las otras dos cepas. Se optimizó la fuente de nitrógeno, la fuente de carbono y el pH del medio, en cultivos de células libres y encapsuladas. La mejor fuente de nitrógeno fue el extracto de levadura llegándose a obtener una actividad de 2.46 UI/mL en células libres y 10.1 UI/mL en células encapsuladas. La mejor fuente de carbono fue la paja de soya (Glycine max) como materia cruda e hidrolizada llegándose a obtener una actividad de 1.9 UI/mL y 2.2 UI/mL respectivamente a un pH de cultivo de 8.
Palabras Clave: xilanasas, inmovilización
ABSTRACT
The thermophilic bacteria that produce enzymes have a big interest in biotechnology, due diverse application in the industry for extreme stability. These biocatalysts are used in the preparation of pulp paper, food processing agroindustrial and hemicellulose conversion to biomass. The purpose of this work is to increase the enzymatic production of the xylanases of thermophilic anaerobic bacteria produced by three strains FT3, 2B and P5 which were isolated from the Bolivian plateau. The strain FT3 was selected with significant xilanolitic activity of 2.0 IU / mL compared with the other strains. The optimization of culture medium was performed in free and encapsulated cell, changing the culture medium concerning nitrogen source, carbon source and pH. The best nitrogen source went yeast extract 2.46 IU / mL with bacteria-free and 10.1 IU/mL in bacteria-encapsulated and best carbon source is the soybean straw (Glycine max) as hydrolyzed material and coming up as raw material to get up to 2.2 IU / mL and 1.9 IU / mL respectively at pH 8.
Key Words: xylanases, immobilization.
INTRODUCCIÓN
El xilano es uno de los polisacáridos más abundantes de la naturaleza su principal componente es la D-xilosa, la cual esta unida por enlaces β-1,4. La estructura del xilano no es homogénea y dependiendo de la fuente natural su estructura varía desde una cadena lineal hasta polisacáridos altamente ramificados1. Forma parte de las hemicelulosas presentes en la matriz amorfa de la pared secundaria de los tejidos lignificados de las plantas leñosas. La degradación del xilano requiere de un complejo de enzimas que actúan de una manera cooperativa para convertir el xilano en sus constituyentes más simples.
Los sistemas enzimáticos microbianos suponen actualmente un importante potencial biotecnológico aplicable en numerosas industrias en el ámbito de las denominadas tecnologías limpias o tecnologías sostenibles2. Las industrias son las más beneficiadas por la aplicación de estas enzimas principalmente en las papeleras, textiles, agrícolas y de alimentos. Las hemicelulasas bacterianas y en especial las xilanasas modifican y transforman la lignocelulosa y otros materiales de la pared celular vegetal.
El objetivo de esta investigación es la optimización de las condiciones de cultivo anaeróbico termófilo de la cepa FT3 para aumentar la producción enzimática de xilanasas en un sistema libre e inmovilizado.
MATERIAL Y MÉTODOS
Microorganismos. Se utilizaron cepas bacterianas anaerobias termófilas aisladas de muestras de lodo de aguas termales obtenidas de la región altiplánica de Chaqui-Bolivia (Tabla 1), mediante técnicas de cultivo anaerobio como el Roller tubes, cultivo bifásico y encapsulación en alginato de calcio3.
Tabla 1. Datos Generales de la Cepa Bacteriana FT3

Medio de cultivo. Las cepas bacterianas FT3, 2B, P5 fueron cultivadas en el medio base mineral 11 el cual esta compuesto de: solución A (NH4Cl 200g/L, NaCl 10g/L, MgCl2*6H2O 10g/L, CaCl2*2H2O 5g/L), solución B (K2HPO4*3H2O 200g/L), solución 2 (FeCl2*4H2O 1.5 g/L, HCl 25% 6.5 mL, H3BO3 60mg/L, CaCl2*6H2O 120mg/L, MnCl2*4H2O 100 mg/L, Na2MoO4*2H2O 25mg/L, NiCl2*6H2O 25mg/L, ZnCl2 70 mg/L, CuCl2*2H2O 15 mg/L), solución 3 (Na2SeO3 3 mg/L, NaOH 0.5g/L), solución 4 (NaHCO3 8.5g/100mL), solución. 7 (Biotina 1mg/L, PABA 5mg/L, Vit. B12 5mg/L, Tiamina 10 mg/L) y la solución Reductora (Na2S*9H2O 2%), utilizando como fuente de carbono el xilano 0.2%, el pH del medio es neutro y la inoculación se realizó según la técnica de Hungate para anaerobios4, se cultivó a una temperatura de 60ºC.
La optimización de la fuente de nitrógeno se realizó bajo el siguiente diseño factorial de cuatro niveles: NH4Cl, NaNO3, Extracto de levadura y urea; tomando en cuenta la concentración de nitrógeno del medio base mineral 11(variación de la solución A) la cual fue la concentración estándar de referencia para el cálculo de los demás compuestos (Tabla 2), a la vez se tomó dos variables: células inmovilizadas y libres.

Equivalencia de los valores de COD (Demanda química de oxígeno) con el equivalente de xilano en el medio de cultivo. Para el cálculo de la cantidad de hidrolizado se realizó una equivalencia. Se tomó 0.2% de xilano (fuente de carbono en el medio 11) como parámetro teórico, al cual se determinó la cantidad de materia orgánica susceptible a ser oxidada mediante la técnica COD. Se obtuvo que 0.2% de xilano requieren de 2107.21 mg de oxígeno molecular para una combustión total hasta dióxido de carbono, agua y calor, a partir de esa relación se obtuvo los siguientes datos para la cantidad de hidrolizado como fuente de carbono requerido para el medio de cultivo (Tabla 3).
Tabla 3. Datos para el cálculo de la concentración de hidrolizado requerido

El efecto del pH en la producción de la enzima xilanasa se estudió realizando un diseño factorial de 6 niveles: pH 10, 9, 8, 7, 6 y 5; dos variables: células inmovilizadas y libres.
Inmovilización de la cepa en perlas de alginato. Esta técnica fue desarrollada como una forma de protección para las bacterias y consiste en mezclar alginato de sodio 1.5% con una suspensión de la cepa FT3 en una relación de 3:1 respectivamente. La suspensión-mezcla debe ser homogénea, después es dispersada mediante goteo con jeringa 27G en una solución de cloruro de calcio 1.5% en el cual se llega a polimerizar en constante agitación por dos horas para permitir la formación de las perlas 5, 6, 7.
Pre-tratamiento de la materia cruda. Se realizó un pre-tratamiento del material crudo con distintos métodos de hidrólisis:
Hidrólisis alcalina. Para este método la materia cruda fue tratada con NaOH 1% y KOH 1%, en una relación de 2:25 y llevada al proceso de autoclave a 121ºC por 15 minutos.
Pre-tratamiento con amonio. En este método la materia cruda fue tratada con NH4 a 29.5% en una relación de 5:50, la mezcla fue dejada a temperatura ambiente por 10 días.
Hidrólisis ácida H2SO4. Este método consiste en tratar la materia cruda con ácido sulfúrico al 1.1% en una relación de 22:2, la mezcla fue llevada a un proceso de autoclave a 121ºC por 15 minutos.
Hidrólisis termal. Este método de hidrólisis consiste en mezclar la materia cruda 2 g con un volumen de agua destilada de 18 mL; la mezcla fue llevada a un proceso de autoclave dos veces a 121ºC por 15 minutos.
En todos los casos se realizó la valoración de COD para calcular la cantidad de materia orgánica oxidable y determinar la concentración a utilizarse en los experimentos de optimización de la fuente de carbono.
Caracterización de las xilanasas. Se realizó la caracterización de los extractos enzimáticos procedentes de la cepa bacteriana FT3. Los cultivos de bacterias se centrifugaron a 10.000 rpm durante 10 minutos; los sobrenadantes obtenidos contienen los extractos enzimáticos.
La influencia de la temperatura en la actividad enzimática se estudió en un rango de 30 ºC a 90 ºC por medición de la velocidad de reacción a pH 6, el efecto de la temperatura en la estabilidad se estudió incubando el crudo enzimático a varias temperaturas de: 30, 40, 50, 60, 70, 80 y 90 ºC por un espacio de 30 minutos.
El efecto del pH en la actividad enzimática se estudió a 80 ºC y se varió el pH del medio de reacción desde 3 hasta 10 consecutivamente; la influencia del pH en la estabilidad de la enzima fue estudiada por la exposición del crudo enzimático a valores de pH de sustrato de 3 (tampón citrato de sodio 0.05 M), 4, 5 (tampón acetato de sodio 0.05 M ), 6, 7, 8 (tampón fosfato de sodio 0.05 M), 9 (tampón cloruro de amonio 0.05 M) y 10 (tampón carbonato de sodio 0.05M) y a 80 ºC durante 30 minutos.
Determinaciones analíticas.
Determinación de xilanasas. Se utilizó el método DNS el cual determina la presencia de grupos carbonilos libres (C=O) de los llamados azúcares reductores. Se basa en la utilización de ácido 3,5-dinitrosalicílico para provocar la oxidación de los azúcares y al mismo tiempo su propia reducción (Figura 1). Un mol de azúcar reaccionará con un mol de ácido 3,5-dinitrosalicílico, dando lugar a una relación estequiométrica que permite conocer la cantidad de azucares reductores presentes en la muestra8.
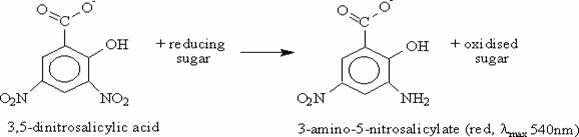
Figura 1. Reacción de oxidación de azucares reductores, Método DNS
Esta reacción es calorimétrica: el ácido 3,5-dinitrosalicílico es de color amarillo, mientras que la aparición del ácido 3-amino, 5-nitrosalicílico provoca un viraje a pardo oscuro-marrón, cuya intensidad será proporcional a la cantidad de azúcares reductores, mostrando de forma indirecta la actividad hidrolítica de la enzima por la cantidad de azúcares reductores presentes producto de la actividad.
Proteínas totales. La determinación de proteínas totales fue realizada por el método de Lowry, usando como estándar albúmina sérica bovina9.
Determinación de biomasa. La biomasa fue determinada por la densidad óptica para el crecimiento microbiano en un espectrofotómetro UV/Vis a una longitud de onda de 560 nm, midiendo la turbidez de éste.
Demanda química de oxígeno COD. El COD mide la porción de materia orgánica biodegradable de una muestra que es susceptible a ser oxidada por un fuerte oxidante químico dicromato de potasio. La muestra se somete a una solución ácida fuerte con un exceso de dicromato potásico. Después de la digestión, el dicromato no reducido sobrante se determina con sulfato ferroso amónico (sal de Morh) (SO4)2Fe(NH4)2 para determinar así la cantidad de dicromato consumido y por lo tanto calcular la materia orgánica oxidable en términos de equivalente de oxígeno.

Ferroína fue utilizada como indicador para la valoración. Ésta reacciona con el exceso de Fe+2 que no ha reaccionado con el dicromato dando lugar a un complejo coloreado que nos indica el punto final de la valoración. (Standar methods, 1992).
RESULTADOS
La selección de la mejor cepa productora de enzimas xilanolítica se realizó en base a los valores de actividad enzimática registrados a través de la determinación de xilanasas por el método DNS. Las condiciones de cultivo empleadas para la selección son: xilano como fuente de carbono y cloruro de amonio como fuente de nitrógeno dando una actividad de 2 UI/mL de la cepa bacteriana FT3 (ver Tabla 4), siendo esta actividad mayor que las otras cepas 2B y P5 cultivadas en las mismas condiciones. Las actividades xilanolíticas fueron de 0.8 para la cepa 2B y 0.64 UI/mL para la cepa FT3. Todos los experimentos fueron realizados por triplicado. Por lo tanto la cepa FT3 fue seleccionada para los siguientes experimentos.
| Tabla 4. Actividades xilanolíticas consideradas para la selección de la mejor cepa termófila anaerobia |

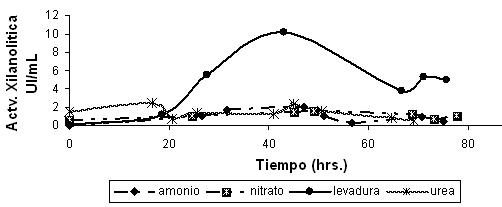
La cepa FT3 fue inoculada en forma libre en medio de cultivo líquido suplementado con xilano como fuente de carbono pero variando la fuente de nitrógeno (Figura 2); registrándose valores de actividad enzimática de 2.5 UI/mL con extracto de levadura existiendo diferencia con los otros compuestos utilizados como fuente de nitrógeno.
|
Figura 2. Efecto de diferentes fuentes de nitrógeno sobre la actividad xilanolítica de la cepa FT3 libre crecida con xilano como fuente de carbono a pH neutro. |

La cepa FT3 también fue inoculada de forma inmovilizada en las mismas condiciones de fuente de nitrógeno y fuente carbono que la cepa libre; registrándose valores óptimos de actividad de 10.1 UI/mL con extracto de levadura (Figura 3); siendo esta mucho mayor a los otros compuestos utilizados como fuente de nitrógeno, motivo por el cual fue seleccionado para los siguientes experimentos (Tabla 5). La encapsulación de la bacteria incrementa la biomasa, lo cual puede evidenciarse por el cambio de la coloración en el centro de la perla pasando de incoloro a un color blanquecino demostrando así crecimiento celular. En los cultivos encapsulados también presentó actividad expresando que las bacterias pueden crecer dentro de las perlas de alginato.
|
Figura 3. Efecto de diferentes fuentes de nitrógeno sobre la actividad xilanolítica de la cepa FT3 inmovilizada, con xilano como fuente de carbono a pH neutro. |
| Tabla 5. Actividad xilanolítica de la bacteria FT3 libres e inmovilizadas |

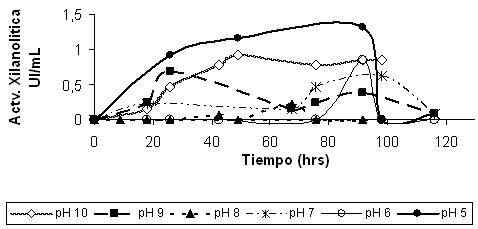
La optimización del pH del medio del cultivo consistió en la variación del mismo, utilizando como fuente de nitrógeno el extracto de levadura 1 g/L, fuente de carbono xilano 0.2% e inoculando a la cepa FT3 en forma inmovilizada. La máxima actividad fue de 1.3 UI/mL a pH 5 (Figura 4) pero no existiendo gran diferencia con los otros pH del medio de cultivo; siendo a la vez que las perlas de alginato son más consistentes por lo cual probablemente provocó la disminución de la porosidad de dichas perlas y la accesibilidad de la bacteria al sustrato. La disminución de la actividad enzimática fue probablemente al cambio de agente reductor de sulfuro de sodio a tioglicolato de sodio 0.05% en el medio de cultivo, ya que por conocimiento el sulfuro de sodio modifica el pH del medio de cultivo debido a los grupos -SH-, a la vez modifica el potencial redox volviéndose el medio aerobio y de esta manera disminuyendo el crecimiento.
|
Figura 4. Efecto de la variación del pH sobre la actividad xilanolítica de la cepa bacteriana FT3 inmovilizada, crecida en xilano como fuente carbono y extracto de levadura como fuente de nitrógeno. |
La cepa FT3 a la vez también fue inoculada en forma libre en las mismas condiciones de cultivo que la cepa inmovilizada, dando como óptimo a pH 8 con una actividad xilanolítica de 3.39 UI/mL; siendo ésta mayor que la actividad a pH ácido (Figura 5), pero menor en comparación con la actividad de la fuente de nitrógeno (Figura 3). Por tal motivo no fue tomado en cuenta este pH del medio de cultivo para los experimentos realizados.
|
Figura 5. Efecto de la variación del pH sobre la actividad xilanolítica de la cepa bacteriana FT3 libre crecida en xilano como fuente carbono y extracto de levadura como fuente de nitrógeno. |

Así también, se optimizó la fuente de carbono utilizando en este caso materia cruda pre-tratada; con el propósito de obtener solamente hemicelulosas10.
La cepa FT3 fue cultivada en medio basal 11 utilizando como fuente de nitrógeno, el extracto de levadura; y como fuente de carbono, hidrolizados de cascarilla de arroz, paja de quinua, paja de trigo, paja de soya. Como control se tomó al xilano; dando una actividad de 2.2 UI/mL con el hidrolizado de paja de soya (Figura 6). Los demás resultados mostraron una actividad similar.
|
Figura 6. Efecto de la variación de las fuentes de carbono (hidrolizados) sobre la actividad xilanolítica de la cepa bacteriana FT3 libre cultivada en extracto de levadura como fuente de nitrógeno a pH neutro. |
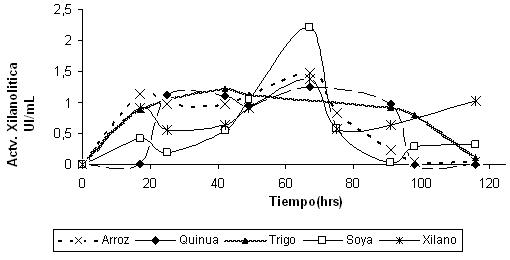
Las diferentes fuentes de carbono como materia pre-tratada no tuvieron gran influencia en la actividad enzimática de las xilanasas, sin embargo se seleccionó al hidrolizado de la paja de soya por presentar una actividad superior a los otros.
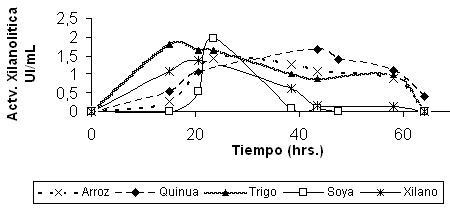
También se realizó la optimización de la fuente de carbono con materia cruda pero solamente con cepas libres y no así con las encapsuladas, debido a la inaccesibilidad que estas tendrían. La cepa bacteriana FT3 pudo crecer con sustratos como paja de soya y paja de trigo presentando una actividad xilanolítica de 1.9 y 1.8 UI/mL respectivamente (Figura 7), siendo éstas inferiores a las actividades presentadas con pre-tratamiento de la materia cruda, probablemente debido a que la enzima no tiene acceso sobre la hemicelulosa.
|
Figura 7. Efecto la variación de las fuentes de carbono (materia cruda) sobre la actividad xilanolítica de la cepa bacteriana FT3 cultivada con extracto de levadura como fuente de nitrógenoa pH neutro. |
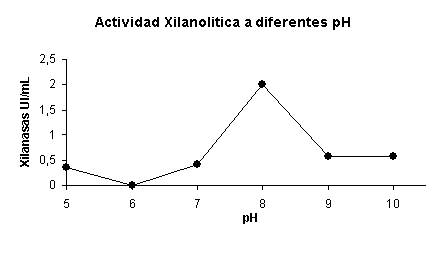
También se realizó la caracterización parcial del extracto crudo enzimático, probando a distintos pH el sustrato. El mismo fue preparado con distintos tampones de acuerdo al pH (tampón citrato, tampón acetato, tampón fosfato, tampón carbonato, tampón de amonio) tomando muestras que se encontraban en sus fases estacionarias, las cuales provenían de un cultivo suplementando con extracto de levadura y con hidrolizado de paja de soya como fuente de carbono y cultivadas a pH neutro. La muestra fue tomada tanto para la actividad de pH y temperatura. La actividad óptima en la reacción enzimática fue a pH 8 dando una actividad xilanolítica de 2 UI/mL (Figura 8).
|
Figura 8. Estabilidad de la enzima a distintos pH de sustrato de la reacción enzimática xilanolítica |
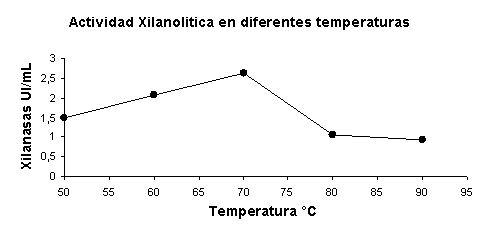
Se realizó a la vez la estabilidad de la enzima a distintas temperaturas de reacción. Fue estable a 70 ºC obteniendo como un máximo de actividad de 2.62 UI/mL (Figura 9).
|
Figura 9. Estabilidad de la enzima a distintas temperaturas de la reacción enzimática xilanolítica |
DISCUSIÓN
La selección de la cepa bacteriana se basó en dos criterios: la producción de la enzima extracelular xilanolítica y el desarrollo en medio de cultivo sólido; teniendo de esta forma la certeza de que se trata de una cepa pura. De las tres cepas bacterianas estudiadas la única que cumplió los dos criterios fue la cepa bacteriana FT3 (bacilo gramnegativo) mostrando una actividad superior a las otras cepas bacterianas 2B y P5, siendo además que no se tuvo la certeza de la pureza de esta dos últimas cepas.
Así también se ha visto que la mejor fuente de nitrógeno en ambos tipos de cultivos libres y encapsulados es el extracto de levadura; probablemente debido a que es un producto rico en vitaminas, aminoácidos y otros factores de crecimiento que serían los responsables de la mayor producción de la enzima xilanasa. Existió también gran diferencia entre ambas variables (inmovilizadas y libres), siendo mejor el sistema inmovilizado. La encapsulación es una forma de proteger el material biológico de factores externos tales como el cambio de pH, metabolitos tóxicos. También la encapsulación incrementa la biomasa y por ende la actividad enzimática lo cual se evidencia por el cambio de coloración dentro de la perla tornándose un color blanquecino.
Por otro lado en la optimización del pH del medio de cultivo se observó que las perlas de alginato son estables a pH ácidos aumentando la consistencia de la perla, además disminuyendo la porosidad de la misma debido al soporte y al pH del medio. Las perlas de alginato no son estables a pH alcalinos llegándose a despolimerizar completamente en el lapso de 2 días en el medio de cultivo, incrementando de esta manera la densidad del medio de cultivo debido a que se entremezcla con el sustrato siendo poco accesible para la bacteria. Por todos los motivos expuestos se decidió trabajar con células libres.
Así también, se ha determinado que para alcanzar mayor nivel de producción de la enzima xilanasa, es necesario dotar a la bacteria de sustratos con pre-tratamiento de la materia cruda (paja de soya); debido a que éstas tienen actividad sobre las hemicelulosas.
En esta investigación la actividad xilanolítica fue incrementada cuando en el medio de cultivo se utilizó como fuente de nitrógeno al extracto de levadura y como fuente carbono el hidrolizado de la materia cruda paja de soya a un pH de 8.
AGRADECIMENTOS
A ASDI/SAREC – Suecia en el desarrollo del Proyecto Biodiversidad Microbiana del Lago Poopó y Río Desaguadero, por el soporte económico.
REFERENCIAS
1. Mayorga L, Gutierrez A, Salgado L, Ponce T. Aislamiento de una clona que contiene un gen de xilanasa a partir de una genoteca de cellulomonas flavigena. Rev Mexicana de Ciencias Farmacéuticas. 2005; 36(2):5-9. [ Links ]
2. Gallardo O. Caracterización de nuevas xilanasas bacterianas.ingeniería de enzimas con la xilanasa B de paenibacillus barcinonensis [tesis doctoral]. Barcelona: Universidad de Barcelona, Facultad de Biología, Departamento de microbiología; 2007. [ Links ]
3. Rios N, Crespo CF, Terrazas LE, Alvarez MT. Aislamiento de cepas anaeróbicas termófilas productoras de celulasas y hemicelulasas implicadas en la producción de Bioetanol mediante técnicas de cultivo y aislamiento tradicionales y no tradicionales. BIOFARBO. 2007; 15 (1): 43-50. [ Links ]
4. Kulasingam T. Cultivating the uncultured: amylolytic microrganisms from ecological niches [Report of thesis Doctoral]. Southern Sweden: Lund University; 2005. [ Links ]
5. Smidsrod O y Skjak B. Alginate as an inmolization matrix for cells. Trends Biotechnol. 1990; 8(3):71-78.
6. Delphine B, Karine J, Thierry L. Optimisation of the cell release from immobilised cells of Bacillus simplex cultivated in culture media enriched with Cd2+: influence of Cd2+, inoculum size, culture medium and alginate beads characteristics. Biotechnol Lett. 2002; 24: 1237–1241. [ Links ]
7. Ivanova E, Chipeva V, Ivanova I, Dousset X, Poncelet D. Encapsulation of lactic acid bacteria in calcium alginate beads for bacteriocin production. J Cult Collect. 2000-2002; 3:53-58. [ Links ]
8. Hansson T, Adlercreutz P. Optimization of Galactoligosaccharide production from lactose using B- glycosidases from Hyperthermophiles. Food Biotechnol. 2001; 2:15. [ Links ]
9. Manual de procedimientos. Universidad de Buenos Aires, Laboratorio de Micología Experimental; 1999. [ Links ]
10. Hendriks ATWM, Zeeman G. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresource Technol. 2009; 100 (1):10–18. [ Links ]