








Serviços Personalizados
Artigo
 Artigo em PDF
Artigo em PDF Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
Links relacionados
 Citado por SciELO
Citado por SciELO
 Similares em SciELO
Similares em SciELO
Bookmark
APTHAPI
versão impressa ISSN 0102-0304
Apthapi v.3 n.3 La Paz 2017
ARTÍCULOS CIENTÍFICOS ORIGINALES
Patogenicidad de Beauveria bassiana sobre Hypothenemus hampei en generaciones reproductivas
Pathogenicity of Beauveria bassiana about Hypothenemus hampei in reproductive generations
Carlos López Blanco *.
* Docente Investigador, Facultad de Agronomía - UMSA. calobla9@.yahoo.es
Recibido: 15/09/2017. Aprobado: 15/11/2017.
Resumen:
El presente estudio se realizó en la Estación Experimental Coroico, San Pedro de la Loma, Municipio de Coroico, durante el año 2013. El objetivo del estudio fue determinar la patogenicidad de cepas de Beauveria bassiana sobre Hypothenemus hampei en 4 cultivos sucesivos. Las variables estudiadas fueron: la conidiación en sustratos y viabilidad de conidias, el grado de patogenicidad y virulencia de las cepas del hongo. Se desarrolló la producción de conidias en sustrato de arroz. La prueba patogénica se realizó en bioensayo en laboratorio, durante 12 días, bajo el Diseño Completamente al Azar con 4 tratamientos (cepas 13, 24, 9205, Testigo), 3 repeticiones, se inoculo a las brocas por inmersión por 3 minutos en una concentración fúngica de 1 x 107 c.ml"1 y el testigo con Agua Destilada Estéril (ADE), confinadas individualmente en viales. Como resultado del estudio, las cepas 9205 y 13 registraron mayor producción de conidias en relación a la cepa 24, en la viabilidad de conidias las cepas no presentaron diferencias significativas superando el 90% de germinación en las diferentes replicas. Referente a la patogenicidad hay diferencias estadísticas significativas en el modo de acción de cepas del hongo sobre la broca, con mortandad alta de las cepas hasta las réplicas 1 y 2, rescatando a la cepa 13 en la réplica 3. La virulencia de cepas manifestó una pérdida gradual en los cultivos sucesivos, con alta agresividad en las réplicas 1 y 2, en términos de mortalidad diaria y tiempo letal medio (TL50), rescatando a la cepa 13 y 9205 en la réplica 3 previa validación, descartando la cepa 24 y la réplica 4 para su aplicación en campo.
Palabras clave: Patogenicidad, virulencia, cepas, conidias, viabilidad.
Abstract:
The present study was carried out at the Coroico Experimental Station, San Pedro de la Loma, municipality of Coroico, during the year 2013. The objective of the study was to determine the pathogenicity of Beauveria bassiana strains on Hypothenemus hampei in four successive crops. The variables studied were conidiation in substrates and viability of conidia, degree of pathogenicity and virulence of fungus strains. The production of conidia in rice substrate was developed. The pathogenic test was performed in a laboratory bioassay for 12 days under the Completely Random Design with 4 treatments (strains 13, 24, 9205, Witness), 3 replicates, inoculated to the drills by immersion for 3 minutes in a concentration 1 x 107 c.ml1 fungicide and the Sterile Distilled Water (ADE) control, individually confined to vials. As a result of the study, strains 9205 and 13showed higher conidia production in relation to strain 24, in conidia viability the strains did not present significant differences, exceeding 90% of germination in the different replicates. Regarding pathogenicity there are significant statistical differences in the mode of action of fungus strains on the borer, with high mortality of the strains up to replicates 1 and 2, rescuing the strain 13 in replícate 3. The virulence of strains showed a gradual loss in the successive cultures, with high aggressiveness in replicates 1 and 2, in terms of daily mortality and mean lethal time (TL50), rescuing strain 13 and 9205 in replícate 3 prior validation, discarding strain 24 and replication 4 for application in the field.
Keywords: Pathogenicity, virulence, strains, conidia, viability.
INTRODUCCIÓN
El hongo entomopatógeno Beauveria bassiana (Balsamo) Vuillemin se encuentra naturalmente infectando a la broca Hypothenemus hampei (Ferrari) en casi todas las regiones donde se encuentre el insecto. Los avances de estudios con el hongo son muy importantes no solo para el control de la broca si no para aplicar a otras situaciones de insectos plaga. Es necesaria la producción del hongo a partir de la selección de aislamientos virulentos, con su reactivación patogénica y manejo del cultivo en sustratos en generaciones, para garantizar la calidad del hongo en términos de patogenicidad y virulencia.
En los Yungas de La Paz se cuenta con cepas promisorias de B.bassiana, para el control de H. hampei, con antecedentes de selección, bioensayo y aplicación. La producción masiva del hongo se realiza a nivel de laboratorio central y comunal (UAC-CP, U-PROAGRO e ILLAMPU) en sustrato de arroz, a partir de cepas madre por generaciones, desconociendo el grado patogénico en dichas replicas productivas. Por consiguiente, el objetivo del trabajo fue estudiar la pérdida de patogenicidad y virulencia del hongo sobre la broca del café, a través del cultivo en sustrato de arroz en generaciones, en laboratorio.
MATERIALES Y MÉTODOS
El presente estudio se realizó en la Estación Experimental Coroico, comunidad San Pedro de la Loma Municipio de Coroico, Provincia Ñor Yungas del departamento de La Paz. Geográficamente ubicado a 16° 12' 17" de Latitud Sur y 67° 41' 52" de Longitud Oeste, a una altura promedio de 1.630 metros sobre el nivel del mar. El método de trabajo se basó en las siguientes fases de estudio:
Reconocimiento y acondicionamiento de laboratorio
Con la recomendación de CENICAFE (1996), se hizo un reconocimiento y acondicionamiento de laboratorio (áreas de preparación, inoculación y maduración) para el proceso productivo del hongo, manejándose bajo condiciones aséptica a una temperatura promedio de 24 °C.
Identificación de cepas del hongo B. bassiana
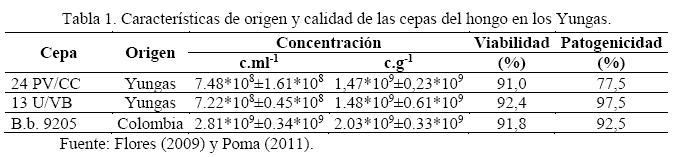
Se identificó las cepas existentes de B. bassiana referente a sus antecedentes de origen, características físicas (Figura 1) y de calidad patogénica en el medio local (Tabla 1).

Reproducción masiva del hongo
- Preparación del sustrato de arroz
Con la recomendación de Poma (2011), se trabajó con arroz cateto, separando elementos indeseables como cascarillas, piedrecillas, semillas de malezas, granos picados. El arroz seleccionado se lavó con agua corriente, quitándose el polvillo, luego se hidrató en agua hervida por 0,60 horas, se escurrió el exceso de agua en un colador (Figura 2), estando el sustrato listo para su embolsado y esterilización.

- Embolsado y esterilización del sustrato de arroz
Para la contención del sustrato se utilizó bolsas de polipropileno, empleando 100 g.bolsa1, cerrando y asegurando con grapas y acomodo en autoclave. Se esterilizó las bolsas por 0,5 h, a una temperatura y presión de 121 °C y 1,5 atmósferas, trasladándose luego a la cámara de flujo laminar, donde se enfriaron y sometieron a luz ultra violeta (acción germicida) por un tiempo de 6 h, para su posterior inoculación.
- Inoculación y maduración del hongo
Se realizó la inoculación sucesiva del hongo a partir de cepas madre (4 generaciones) en sustrato esterilizado en la cámara de flujo laminar con la ayuda del asa de platino y mechero bunsen, registrando las fechas y códigos. Las bolsas inoculadas se agitaron para diseminar las conidias, disponiéndose en la cámara de incubación, donde se hizo un seguimiento de ±20 días, evidenciando su maduración.
Obtención de H. hampei para bioensayos
Con el fin de obtener brocas de una misma edad para los bioensayos de patogenicidad, se multiplico la broca en laboratorio sobre granos pergamino (45% de humedad), a partir de brocas recolectados de campo. Se desinfecto las brocas con hipoclorito de sodio al 0,5%, lavados con Agua Destilada Estéril (ADE) y secados con papel toalla, eligiendo las brocas activas para su infestación en grano pergamino.
Lecturas de concentración y dosificación de conidias del hongo
Con la metodología de Antia et al. (1992), el conteo de conidias del hongo se realizó posterior a los 25 días. Se preparó la solución madre o dilución 10° con la mezcla del hongo (100 g), ADE (200 mi) y aceite agrícola (0,1%). Asimismo, se preparó tubos de ensayo con 9 mi de ADE en orden secuencial en gradilla. Se tomó de la solución madre previa agitación la submuestra de 1 mi depositando al primer tubo, logrando la dilución 101, y de este 1 mi al siguiente tubo, logrando la dilución 102 y así sucesivamente, logrando la dilución adecuada en 104, con la ayuda del microscopio y la cámara de Neubauer, lo que permitió el conteo de conidias 10 a 50 conidias por cuadrante. La cámara de Neubauer tiene 2 retículos, cada una subdividida en 9 cuadros de 1 mm2 con un área total de 9 mm2, para el caso se utilizó el cuadrante central subdividido en 25 cuadrantes y estos en 16 cuadros más pequeños, cuyo factores 104.
La dilución adecuada (104) se agito y se tomó muestras de 0,01 mi, depositando en cada retículo de la cámara; con el microscopio (objetivo 10X y 40X) se procedió con el conteo de conidias (Figura 3). La concentración de conidias se calculó empleando la formula siguiente:
C = N * Dilución empleada * Factor cámara
Dónde:
C: Concentración que se desea conocer (c.ml-1)
N: Número promedio de conidias por cuadrante.

- Solución madre Dilución 10°; Diluciones (ÍO^IO2, 103, 104) a partir de 10°; Cámara Neubauer; Localización de campos de conteo con microscopio (10X, 40X); Retículo central de conteo de la cámara Neubauer.
En base a CENICAFE (1996), el conteo se estableció sumando el total de conidias presentes en los 25 cuadrantes centrales de la cámara, realizando 3 veces este procedimiento con 6 lecturas. Asimismo, en base a González et al. (1993); Bernal et al. (1994), se ajustó la dosis de cepas del hongo a una concentración de 1*107 c.ml-1 para la inoculación de brocas, asemejando a las condiciones de campo.
Prueba de viabilidad del hongo
En base a Marín (1994), se preparó el cultivo Saboraud Dextrosa Agar (SDA) en cajas Petri, donde se inoculo las cepas del hongo a partir de submuestras de la dilución 104, a razón de 5 alícuotas, incubado por 12 h a 24 °C. Se procedió a cortar y fijar las alícuotas en porta objetos con alcohol etílico, usando colorante policrómico para el teñido de conidias. El conteo de germinación de conidias se hizo en microscopio, contando 100 conidias por alícuota, calculando el porcentaje de germinación en las réplicas.
Bioensayo de patogenicidad del hongo
- Preparación de viales
Se realizó la esterilización de viales, cortando discos de papel absorbente para la base dentro el vial para captar y mantener la humedad, se preparó motas de algodón (tapón) e identifico los tratamientos.
- Infección y disposición de brocas
Previa selección y desinfección, las brocas se infectaron por inmersión por 3 minutos con la solución ajustada del hongo (107 c.ml1), y las brocas testigo se inocularon con ADE, disponiéndose de forma individual en viales y tapándose con las motas de algodón. A las 24 horas, se adiciono un grano de café pergamino por vial, para la alimentación de las brocas. Asimismo, se adiciono diariamente 0,2 mi de ADE por vial, para mantener la humedad requerida.
Méndez (1990), sostiene que la individualización de brocas permite establecer el efecto directo del hongo, evitando el contagio cuando se evalúan poblaciones, en las que una broca infectada puede enfermar a otras de la población provocando sesgo.
- Evaluación de patogenicidad y virulencia del hongo
En base a Alves (1986) y González et al. (1993), se evaluó la patogenicidad y virulencia de las cepas del hongo sobre la broca del café en 4 bioensayos consecutivos, por 12 días, registrando los tiempos de mortandad por causas patogénicas y el ciclo del hongo, manteniendo las brocas vivas o muertas en los viales para no interrumpir la dinámica del insecto y el desarrollo hongo.
CENICAFE (1996), señala que el bioensayo determina la ocurrencia de mortandad por causas patogénicas, por otras causas y sobrevivencia, considerando el 100% de la población de insectos.
Variables de estudio
- Concentración y viabilidad promedio de cepas del hongo en 4 réplicas reproductivas sucesivas, mediante la esporulación (c.ml1) y germinación (%) de conidias en tiempos determinados.
- La patogenicidad de las cepas del hongo sobre la población de brocas en las respectivas replicas (Mortandad por el hongo, otras causas y sobrevivencia de brocas).
- El grado de virulencia del hongo sobre la broca del café en términos de mortalidad diaria, mortalidad acumulada, Tiempo Letal Medio (TL50) y tiempos en las etapas del ciclo de desarrollo.
Diseño experimental
El estudio se llevó bajo el diseño "Completamente al Azar" con 4 tratamientos, 3 repeticiones, empleando 10 brocas por tratamiento, con un margen de error del 5%.Se realizó las pruebas: Análisis de Varianza, Pruebas Duncan, Desviación Estándar y Límite de Confianza (Little y Hills, 1990). Los datos obtenidos (porcentaje) se sometieron a transformación angular (Grados) (arcseno √X).
RESULTADOS Y DISCUSIÓN
Concentración y viabilidad de conidias del hongo
La esporulación promedio del hongo en 4 réplicas fue: 2,83*108 ± 0,13*108; 5,51*108 ± 0,45*108; 5,93*108± 0,74*108 c.ml-1 en las cepas 24, 13 y 9205, con superioridad de la cepa 9205 y 13, respecto a la cepa 24, indicando la variabilidad genética de cepas en conidiación. La viabilidad del hongo, hasta 12 h de incubación, manifestó porcentajes de germinación promedio de: 91,30 ± 0,68%, 91,69 ± 1,54% y 91,20 ± 1,66% en las cepas 24, 13 y 9205; superando el parámetro establecido de 85% (Marín, 1994).

- Producción masiva del hongo en sustrato de arroz; conidiación; retículo con conidias; germinación de conidias.
Alves (1986), indica que los hongos entomopatógenos presentan una variabilidad genética recurrente de heterocariosis que resulta con diferentes grados de patogenicidad, virulencia, especificidad, producción de conidias y resistencia a factores abióticos. La calidad del inoculo de un entomopatógeno se relaciona con la viabilidad de conidias generando variabilidad en las respuestas de patogenicidad sobre el hospedero (Lazo, 1990).
Evaluación de patogenicidad del hongo sobre la broca
Se observó en brocas tratadas con el hongo los síntomas de enfermedad, reflejado en la dinámica del insecto, con menor actividad en brocado de granos, con pérdida de coordinación en sus movimientos en relación al testigo que manifestó una actividad dinámica.
En este sentido Alves (1986), insectos adultos infectados por B. bassiana presentan movimientos lentos, pierden coordinación, no se alimentan, reducen su radio de acción y son afectados en la ovoposición.
- Análisis de Varianza de patogenicidad en cultivos sucesivos
El Análisis de Varianza de patogenicidad de cepas de B. bassiana en 4 réplicas reproductivas, presentan diferencias altamente significativas, indicando que el desenvolvimiento patogénico de cepas del hongo sobre la broca del café, es de forma diferenciada (Tablas 2, 3, 4 y 5).
Con la prueba Duncan (Tablas 6, 7, 8 y 9), se comparó estadísticamente la patogenicidad del hongo sobre la broca (mortandad promedio), con los siguientes resultados: Replica 1, similitud y superioridad patogénica de las cepas 13 y 9205 respecto a la cepa 24. Replica 2, similitud patogénica de cepas. Replica 3, superioridad de la cepa 13 respecto a las otras 2 cepas. Replica 4, similitud patogénica de cepas. El testigo en las respectivas réplicas no presentó mortandad por causas patogénicas.
Se evidencia la pérdida gradual de patogenicidad de cepas del hongo con descenso de mortandad de brocas en las réplicas reproductivas.
- Mortandad y sobrevivencia de la broca del café
La ocurrencia de mortalidad por causas patogénicas del hongo, mortalidad por otras causas y la sobrevivencia de brocas se detallan en el Cuadro 10. Los datos evaluados sobre 30 individuos, representan el 100%, lo que equivale a que cada broca y/o individuo tome el valor de 3,33%.
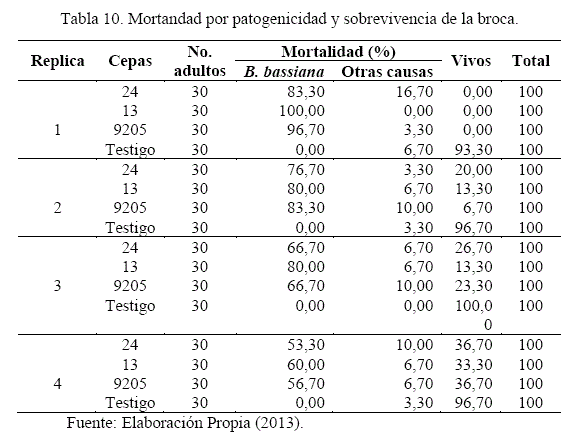
La mortandad de brocas por efecto patogénico del hongo manifestó un descenso progresivo en las réplicas reproductivas, con alta patogenicidad de las cepas del hongo estudiadas (13, 24 y 9205) en las réplicas 1 y 2, y recuperando la cepa 13 en la réplica 3 (recomendados para su aplicación en campo).
En la mortandad por otras causas, en las réplicas con cepas del hongo, presentan rangos desde 16,70% a 3,30%, con posibles causas de mortandad por manipuleo de granos, aplastamiento y otros.
Referente a la sobrevivencia de brocas, las tratadas con el hongo, manifiestan un 100% de mortalidad en la réplica 1, con sobrevivencia de insectos en las réplicas siguientes, de 6,70% a 36,70%, indicando la pérdida gradual de patogenicidad de cepas del hongo en los pases inoculativos. El testigo presentó más de 90% de sobrevivencia, confirmando el efecto patogénico del hongo, en mayor y/o menor grado.
Kawakami (1960), Schaerffenberg (1964), Fargues (1972), Nagaich (1973), Hall (1980), Morrow et al. (1989), Ignoffo etal. (1992); citado por González et al. (2001), indican que los hongos entomopatógenos cultivados en medios no enriquecidos o carentes de pases en insectos, reducen su patogenicidad en pases sucesivos, demostrado con los hongos B. bassiana, Metarhizium anisopliae y Verticillium lecanii.
Grado de virulencia del hongo
- Mortandad diaria de población de brocas
La mortalidad diaria de población de brocas por acción de las cepas del hongo en las respectivas réplicas de cultivos (Figura 5), presentan diferentes distribuciones durante los periodos evaluados.

El análisis de los resultados de mortalidad diaria de brocas por causas patogénicas de las cepas del hongo en las réplicas reproductivas son las siguientes:
Replica 1, la mortandad en la cepa 24, se inicia después de 48 horas (50,9 h), con mayor mortandad (44%) hasta las 60 horas, finalizando después de 132 horas (136,8 h). En la cepa 13, se inicia antes de las 24 horas (20,3 h), con mayor mortandad (43,3%) hasta las 60 horas, finalizando antes de 108 horas (104,9 h). En la cepa 9205 se inicia después de 36 horas (41,9 h), con máxima mortandad (24,1%) hasta 72 horas, finalizando después de 132 horas (136,8 h). Los resultados indican un alto grado de virulencia de las cepas del hongo, con leve superioridad de la cepa 13, respecto a las cepas 24 y 9205.
Replica 2, la mortalidad con la cepa 24 se inició después de 48 horas (55,8 h), con mayor mortandad (21,7%) hasta 96 horas, finalizando antes de 180 horas (173,2 h). En la cepa 13, se inicia a las 48 horas, con máxima mortandad (33,3%) hasta 72 horas, finalizando antes de 120 horas (118,5 h). En la cepa 9205 se inicia a las 48 horas, con la máxima mortandad (28%) hasta 84 horas, con extensión antes de 144 horas (140,7 h). Los resultados indican la pérdida de virulencia de cepas del hongo con relación a la réplica 1, siendo superior la cepa 13 en relación a las cepas 9205 y 24.
Replica 3, la mortandad con la cepa 24 se inicia después de 48 horas (51,5 h), con máxima mortandad (25%) hasta 96 horas, finalizando después de 168 horas (170,8 h). La cepa 13, inicia la mortandad a las 48 horas, con máxima mortandad (29,2%) hasta 72 horas, finalizando después de 144 horas (148,8 h). La cepa 9205, inicia a las 48 horas, con la máxima mortandad (20%) a 72 y 84 horas, finalizando después de 144 horas (148,8 h). Los resultados indican la perdida de virulencia significativa, en especial de la cepa 24, rescatando las cepas 13 y 9205 para su aplicación en campo (previa valoración).
Replica 4, con la cepa 24, la mortandad se inicia después de 84 horas (88,7 h), con mayor mortandad (25%) hasta 132 horas, finalizando después de 168 horas (171,8 h). En la cepa 13, se inicia después de 72 horas (74,8 h), con máxima mortandad (26,3%) hasta 108 horas, finalizando después de 144 horas (149,4 h). En la cepa 9205 se inicia, igual que en la anterior, con máxima mortandad (23,5%) a las 96 horas, finalizando después de 156 horas (159,6 h). Los resultados en esta fase reproductiva, indican una alta pérdida de virulencia, con baja efectividad de control de poblaciones de broca de café.
- Tiempo Letal Medio (TL50)
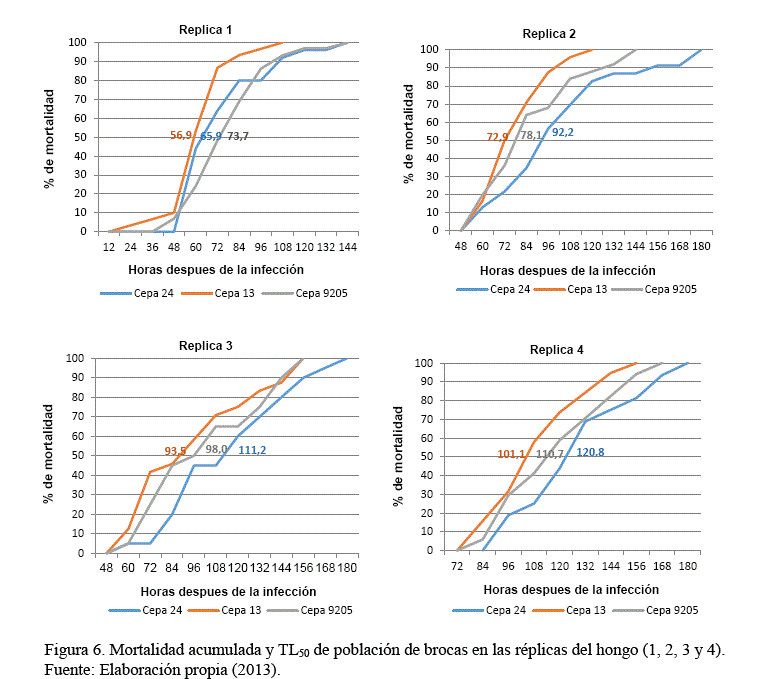
La Figura 6 (gráficas), muestra el comportamiento variable de la mortalidad acumulada (100%) y la mortalidad del 50% (TL50) de población de brocas, por efecto del grado patogénico de cepas del hongo en las réplicas reproductivas, cuyo análisis de resultados se detalla a continuación:
Replica 1, la cepa 13 fue superior con un Tiempo Letal medio (TL50) de 56,9 horas (2,4 días) respecto a las cepas 24 y 9205 que presentaron 65,9 (2,8 días) y 73,7 horas (3,1 días). Los resultados manifiestan la alta virulencia de las cepas evaluadas sobre la población de brocas, garantizando su aplicación y efectividad bajo condiciones de campo.
Replica 2, superioridad de la cepa 13, con TL50 de 72,9 horas (3 días), respecto a las cepas 9205 y 24 con tiempos de 78,1 y 92,2 horas (3,3 y 3,8 días). Se tiene la pérdida de virulencia de las cepas del hongo en relación a la réplica 1, siendo aun altamente virulentos para el control de la broca en campo.
Replica 3, los TL50, de mortalidad de brocas indican tiempos de 93,5 horas (3,9 días), 98 horas (4,1 días) y 111,2 horas (4,6 días) en las cepas 13, 9205 y 24. Existe una pérdida de virulencia significativa con relación a las réplicas 1 y 2, rescatando a la cepa 13 y 9205 (TL50 aceptable), para su aplicación en campo (previa validación), se descarta a la cepa 24 en esta fase reproductiva por su alto valor en TL50.
Replica 4, los TL50, de cepas del hongo presentan tiempos mayor a 100 horas, indicando la afectación de la virulencia en mayor proporción en relación a las anteriores replicas, debiendo descartar su aplicación en campo. Se recomienda reactivar las cepas del hongo sobre la broca para recuperar su virulencia.
La pérdida de virulencia de las cepas del hongo sobre la broca del café en las réplicas es evidente, atribuyendo a factores fisiológicos y genéticos de las cepas evaluadas.
Al ves (1986), indica que un patógeno de alta virulencia incide sobre un gran número de individuos produciendo epizootias con gran capacidad de diseminación, siendo seleccionado para el control microbiano de plagas. A su vez CENICAFE (1995), señala que aislamientos de hongos obtenidos directamente de insectos son más virulentos, permitiendo obtener cultivos de primera generación.
Fargues y Roberts (1983); citado por González et al. (2001), señalan que cultivos sucesivos de entomopatógenos resultan en una pérdida de la virulencia (CL5o, TL5o), debiendo evitar estos problemas con la producción de grandes cantidades de inoculo inicial y almacenarlo. La pérdida de virulencia depende del aislamiento y especie del patógeno, recuperando su agresividad con el pasaje del hongo a través del hospedero apropiado.
A su vez Bustillo (2002), señala que cuando se cultiva el hongo en sustratos por tres o más generaciones su patogenicidad se reduce considerablemente y el tiempo promedio para causar mortalidad en la mitad de la población se incrementa, en comparación con el hongo activado sobre la broca (reactivación patogénica).
Asimismo, Brownbridge et al. (2001); citado por García et al. (2006), atribuyen el efecto diferencial de los cultivos sucesivos en la patogenicidad de patógenos a una variación inter e intra específica de estabilidad genética, por las diferencias en metodologías usadas, mutaciones y el efecto de la condición de cultivos.
- Tiempos en las etapas de desarrollo del hongo
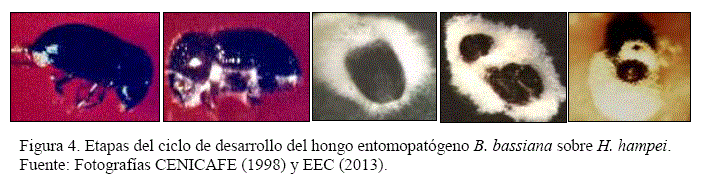
El ciclo de desarrollo del hongo sobre la broca presentó la fase patogénica y saprofítica, con la muerte del insecto por acción enzimática y toxica del hongo en el primer caso, y con la formación de estructuras del hongo en el segundo caso, con las etapas de producción de micelio, cubrimiento micelial, conidiogénesis y liberación de conidias (Figura 4). Los tiempos en las diferentes etapas de desarrollo del hongo se detalla en el Cuadro 11, medidos a partir de la inoculación y muerte del insecto en las fases citadas.
- Etapas de desarrollo: muerte, producción de micelio, cubrimiento micelial, conidiogénesis y liberación de conidias.
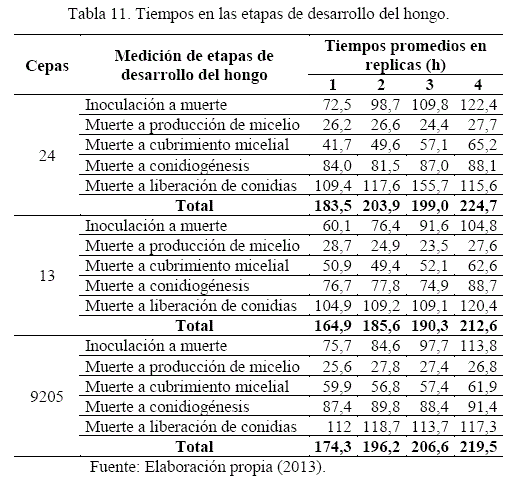
El análisis de etapas de desarrollo del hongo en base al cuadro anterior se describe a continuación:
Inoculación a muerte, etapa con tiempos variables de mortandad, siendo superior la cepa 13 con tiempos de 60,1 a 104,8 horas en relación a las cepas 9205 y 24 con tiempos después de 70, 110 y 120 horas, evidenciando la pérdida del grado patogénico y virulento de cepas del hongo en las réplicas sucesivas.
Muerte a cubrimiento micelial, de la misma manera existe poca variabilidad de tiempos en las cepas del hongo sobre la broca de café, obteniendo un promedio de 55,4 horas.
Muerte a conidiogénesis total, con leve superioridad de la cepa 13 con un tiempo de 79,5 horas respecto a las cepas 24 y 9205 con tiempos de 85,2 y 89,2 horas.
Muerte a liberación de conidias, con superioridad de la cepa 13, con 110,9 horas respecto a las cepas 9205 y 24 con 115,4 y 124,6 horas, con conidiación pulverulenta.
El Tiempo total del ciclo en las cepas del hongo sobre la broca de café en las réplicas respectivas, tiene un ascenso gradual, ratificando la afectación del hongo con pérdida de patogenicidad y virulencia.
En este sentido García et al. (2006), afirman que la actividad insecticida de algunos aislamientos de hongos entomopatógenos, depende de las características genéticas, el origen geográfico, el hospedero de origen, capacidad de esporulación, actividad enzimática y de la viabilidad de conidias.
CONCLUSIONES
La viabilidad de cepas de B. bassiana, realizado en Saboraud Dextrosa Agar (SDA), presento una germinación promedio de conidias mayor a 90% en las réplicas (12 horas), superando el parámetro mínimo de comparación exigido de 85%.No influyendo en la patogenicidad de las cepas evaluadas.
El Análisis de Varianza de patogenicidad de B. bassiana expresada con la mortandad de H. hampei, presentó diferencias altamente significativas (P=0,05), indicando la acción patogénica heterogénea de las cepas del hongo sobre la broca del café en las diferentes replicas reproductivas.
Muerte a producción de micelio, etapa con variaciones poco significativas en tiempos, obteniendo un promedio de 26,4 horas en las réplicas reproductivas.
La prueba Duncan, determino estadísticamente la patogenicidad de cepas del hongo sobre población de brocas en las réplicas, con los siguientes resultados: replica 1, grado patogénico similar y superior de las cepas 13 y 9205 respecto a la cepa 24; Replica 2, patogenicidad similar de cepas; Replica 3, la cepa 13 superior a la similitud de las cepas 9205 y 24; Replica 4, patogenicidad similar de las cepas.
La mortandad y sobrevivencia de brocas evidenció la perdida patogénica gradual de cepas del hongo, con mortandad promedio de 93,3%, 80%, 71,1% y 56,7%; sobrevivencia de 0%, 13,3%, 21,1% y 35,6% en las réplicas 1, 2, 3, y 4. Las brocas testigo (sin inoculo) sobrevivieron en 96,7%.
La mortandad diaria de brocas por acción patogénica manifestó la pérdida gradual de virulencia de cepas del hongo en las réplicas, presentando el inicio de mortandad hasta 12, 48 y 76 horas, con máxima mortandad de 44%, 33,3%, 29,2% y 26,3% hasta las 60, 72, 84 y 120 horas extendiéndose hasta las 120, 132 y 168 horas en las cepas 24, 13 y 9205 y replicas 1, 2, 3 y 4 respectivamente.
El tiempo en el cual ocurrió el 50% de mortandad (TL50) de población de brocas por la acción virulenta de cepas del hongo en las réplicas, presentó tiempos promedios de 65,2; 81,1; 100,9 y 110,9 horas en las réplicas 1, 2, 3 y 4, empleando mayor tiempo de control de la plaga en los pases en cultivos.
El ciclo de B. bassiana sobre H. hampei presento la fase patogénica y saprofítica; con las etapas de muerte, crecimiento micelial, cubrimiento micelial, conidiogénesis y liberación de conidias. En las etapas de las fases respectivas se evidencia la perdida de virulencia de cepas del hongo de forma gradual, con superioridad de la cepa 13 con menores tiempos respecto a las cepas 24 y 9205
De manera general se concluye, que los cultivos sucesivos afectaron la patogenicidad y virulencia efectiva de aislamientos de cepas de B. bassiana de manera gradual suponiendo la afectación de sus características morfológicas y fisiológicas, manteniendo su alta calidad agresiva hasta la réplica 2, garantizando su aplicación en campo para el control de la plaga broca de café.
REFERENCIAS BIBLIOGRÁFICAS
Alves, S. B. (1986). Controle microbiano de insectos fungos entomopatogenicos. Sao Paulo, Brasil. Editora Manóle, p. 73 - 75 [ Links ]
Antia, O. P.; Posada, F. J.; Bustillo, A. E.; Gonzales, M. T. (1992). Producción en finca del hongo Beauveria bassiana para el control de la broca del café. CENICAFE. N° 182: 1 - 12. [ Links ]
Bernal, M. G.; Bustillo, A. E.; Posada F., F.J. (1994). Virulencia de aislamientos de Metarhizium anisopliae y su eficacia en campo sobre Hypothenemus hampei. Revista Colombiana de Entomología. 20 (4): 225- 228. [ Links ]
Bustillo, P., A. E. (2002). El manejo de cafetales y su relación con el control de la broca del café en Colombia. Programa de Investigación Científica CENICAFE (Colombia). N° 24: 1- 40 [ Links ]
CENTRO NACIONAL DE INVESTIGACIONES DEL CAFÉ. (1996). Práctica sobre el control de calidad a formulaciones de hongos entomopatógenos In: Seminario Internacional sobre Caficultura sostenible. "Manejo integrado de la broca del café "MIB. Chinchiná, Caldas. Abril 17-19: 33. [ Links ]
Flores, CH., C. R. 2009. Evaluación de patogenicidad de cepas del hongo Beauveria bassiana (Balsamo) Vuill. Sobre la broca del café Hypothenemus hampei (Ferrari) en laboratorio. Universidad Mayor de San Andrés, Facultad de Agronomía. Tesis de Grado. La Paz - Bolivia. P. 62. [ Links ]
GARCÍA, M.; VILLAMIZAR, L.; TORRES, L.; CORTES, A. 2006. Efecto de subcultivos sucesivos de Beauveria bassiana sobre sus características y actividad contra Premnotrypes vorax. Manejo Integrado de Plagas y Agroecología (Costa Rica) No. 77. p 50-56. [ Links ]
González, G., M. T.; Posada, F. F.; Bustillo, P., A. E. (1993). Desarrollo de un bioensayo para evaluar la patogenicidad de Beauveria bassiana sobre Hypothenemus hampei. Revista CENICAFE, (Colombia). 44(3): 93-102. [ Links ]
González, G., M. T.; Valencia, J., A.; Bustillo, P., A. E. (2001). Incremento de la patogenicidad de Beauveria bassiana sobre Hypothenemus hampei, utilizando integumento del insecto en el medio de cultivo. Manejo Integrado de Plagas (Costa Rica) No. 60 p. 31-35 [ Links ]
Lazo, A., R. R (1990). Susceptibilidad de la broca del fruto del cafeto Hypothenemus hampei al hongo entomopatógeno Beauveria bassiana y su tolerancia al oxicloruro de cobre. Turrialba. (Costa Rica). Centro Agronómico Tropical de Investigación y Enseñanza CATIE. 61 p. [ Links ]
Little, T., M.; Hills, F., J. (1990). Métodos estadísticos para la investigación en la agricultura. 2da. Ed. Trillas. México D. F. 125 - 143 p. [ Links ]
Marín, P. (1994). Evaluación de diferentes medios de cultivo para determinar la germinación del hongo Beauveria bassiana producido en formulaciones comerciales. Informe Anual de Labores disciplina de Entomología. CENICAFE. 26 P. [ Links ]
Méndez, L. I. (1990). Control microbiano de la broca del fruto del cafeto Hypothenemus hampei Coleóptera: Scolytidae) con el hongo Beauveria bassiana (Bals.) Vull. (Deuteromycetes) en el Soconusco. Chiapas, Chapingo (México). Colegio de Postgraduados (Tesis Maestría). 135p. [ Links ]
Poma, L., H. J. (2011). Reproducción masiva de conidias de cepas del hongo Beauveria bassiana (Balsamo) Vuill. sobre sustrato de arroz, bajo diferentes calidades de grano. Universidad Mayor de San Andrés. Tesis de Grado para optar al título de Ingeniero Agrónomo. La Paz, Bolivia. 63 p. [ Links ]
ANEXOS